СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Рассматриваемые ниже свойства нервных центров связаны с некоторыми особенностями распространения возбуждения в ЦНС, особыми свойствами химических синапсов и свойствами мембран нервных клеток. Основными свойствами нервных центров являются следующие.
А. Инерционность — сравнительно медленное возникновение возбуждения всего комплекса нейронов центра при поступлении к нему импульсов и медленное исчезновение возбуждения нейронов центра после прекращения входной импульсации. Инерционность центров связана с суммацией возбуждения и последействием.
Явление суммации возбуждения в ЦНС открыл И. М. Сеченов (1868) в опыте на лягушке: раздражение конечности лягушки слабыми редкими импульсами не вызывает реакции, а более частые раздражения такими же слабыми импульсами сопровождаются ответной реакцией - лягушка совершает прыжок. Различают временную (последовательную) и пространственную суммацию (рис. 5.9).
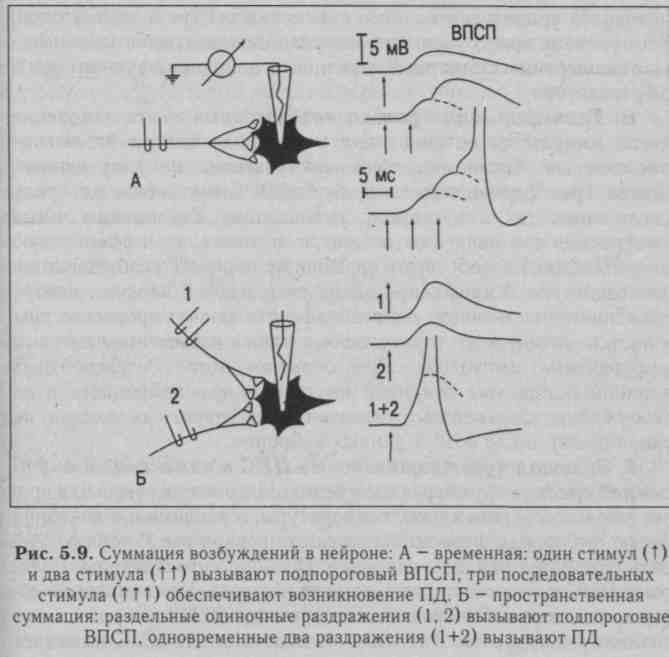
Последействие — это продолжение возбуждения нервного центра после прекращения поступления к нему импульсов по афферентным нервным путям. Основной причиной последействия является циркуляция возбуждения по замкнутым нейронным цепям (см. рис. 5.8), которая может продолжаться минуты и даже часы.
Б. Фоновая активность нервных центров (тонус) объясняется: 1) спонтанной активностью нейронов ЦНС; 2) гуморальными влияниями биологически активных веществ (метаболиты, гормоны, медиаторы и др.), циркулирующих в крови и влияющих на возбудимость нейронов; 3) афферентной импульсацией от различных рефлексогенных зон; 4) суммацией миниатюрных потенциалов, возникающих в результате спонтанного выделения квантов медиатора из аксонов, образующих синапсы на нейронах; 5) циркуляцией возбуждения в ЦНС. Значениефоновой активности нервных центров заключается в обеспечении некоторого
исходного уровня деятельного состояния центра и эффекторов. Этот уровень может увеличиваться или уменьшаться в зависимости от колебаний суммарной активности нейронов нервного центра-регулятора.
В. Трансформация ритма возбуждения — это изменение числа импульсов, возникающих в нейронах центра на выходе относительно числа импульсов, поступающих на вход данного центра. Трансформация ритма возбуждения возможна как в сторону увеличения, так и в сторону уменьшения. Увеличению числа импульсов, возникающих в центре в ответ на афферентную импульсацию, способствуют иррадиация процесса возбуждения и последействие. Уменьшение числа импульсов в нервном центре объясняется снижением его возбудимости за счет процессов пре-и постсинаптического торможения, а также избыточным потоком афферентных импульсов. При большом потоке афферентных влияний, когда уже все нейроны центра или нейронного пула возбуждены, дальнейшее увеличение афферентных входов не увеличивает число возбужденных нейронов.
Г. Большая чувствительность ЦНС к изменениям внутренней среды, например, к изменению содержания глюкозы в крови, газового состава крови, температуры, к вводимым с лечебной целью различным фармакологическим препаратам. В первую очередь реагируют синапсы нейронов. Особенно чувствительны нейроны ЦНС к недостатку глюкозы и кислорода. При снижении содержания глюкозы в 2 раза ниже нормы (до 50% от нормы) могут возникнуть судороги. Тяжелые последствия для ЦНС вызывает недостаток кислорода в крови. Прекращение кровотока всего лишь на 10 с приводит к очевидным нарушениям функций мозга, человек теряет сознание. Прекращение кровотока на 8-12 мин вызывает необратимые нарушения деятельности мозга - погибают многие нейроны, в первую очередь корковые, что ведет к тяжелым последствиям.
Д. Пластичность нервных центров — способность нервных элементов к перестройке функциональных свойств. Основные проявления пластичности следующие.
1. Синаптическое облегчение - это улучшение проведения в синапсах после короткого раздражения афферентных путей. Степень выраженности облегчения возрастает с увеличением частоты импульсов, оно максимально, когда импульсы поступают с интервалом в несколько миллисекунд.
Длительность синаптического облегчения зависит от свойств синапса и характера раздражения - после одиночных стимулов оно невелико, после раздражающей серии облегчение в ЦНС может
продолжаться от нескольких минут до нескольких часов. По-видимому, главной причиной возникновения синаптического облегчения является накопление Са2+ в пресинаптических окончаниях, поскольку Са2+, который входит в нервное окончание во время ПД, накапливается там, так как ионная помпа не успевает выводить его из нервного окончания. Соответственно увеличивается высвобождение медиатора при возникновении каждого импульса в нервном окончании, возрастает ВПСП. Кроме того, при частом использовании синапсов ускоряется синтез рецепторов и медиатора и ускоряется мобилизация пузырьков медиатора, напротив, при редком использовании синапсов синтез медиаторов уменьшается - важнейшее свойство ЦНС. Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах. Значение синаптического облегчения заключается в том, что оно создает предпосылки улучшения процессов переработки информации на нейронах нервных центров, что крайне важно, например, для обучения в ходе выработки двигательных навыков, условных рефлексов.
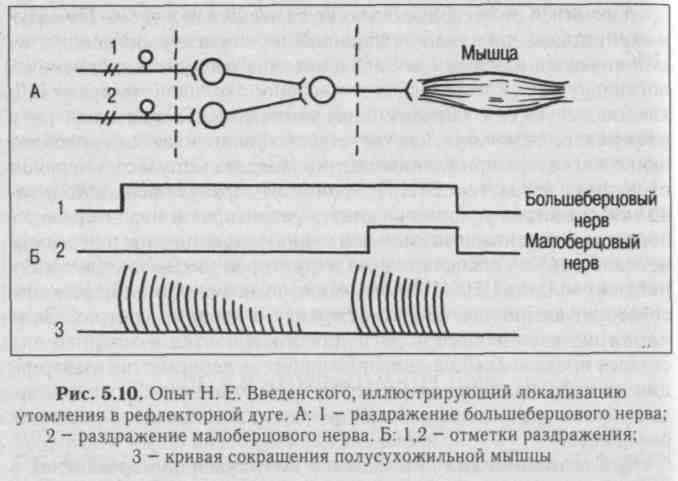
2. Синаптическая депрессия - это ухудшение проведения в синапсах в результате длительной посылки импульсов, например, при длительном раздражении афферентного нерва (утомляемость центра). Утомляемость нервных центров продемонстрировал Н. Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения п. тлЫаНз и п. регопеиз. В этом случае ритмическое раздражение одного нерва вызывает ритмические сокращения мышцы, приводящие к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги (рис. 5.10). Ослабление реакции центра на афферентные импульсы выражается в снижении постси-наптических потенциалов. Оно объясняется расходованием медиатора, накоплением метаболитов, в частности, закислением среды при длительном проведении возбуждения по одним и тем же нейронным цепям.
3. Доминанта - стойкий господствующий очаг возбуждения в ЦНС, подчиняющий себе функции других нервных центров. Доминанта - это более стойкий феномен облегчения. Явление доминанты открыл А. А. Ухтомский (1923) в опытах с раздражением двигательных зон большого мозга и наблюдением сгибания конечности животного. Как выяснилось, если раздражать корковую двигательную зону на фоне избыточного повышения возбудимости другого
4—247
нервного центра, сгибания конечности может не произойти. Вместо^ сгибания конечности раздражение двигательной зоны вызывает реакцию тех эффекторов, деятельность которых контролируется господствующим, т. е. доминирующим в данный момент в ЦНС, нервным центром.
Доминантный очаг возбуждения обладает рядом особых свойств, главными из которых являются следующие: инерционность, стойкость, повышенная возбудимость, способность «притягивать» к себе ирра-диирующие по ЦНС возбуждения, способность оказывать.угнетающие влияния на центры-конкуренты и другие нервные центры.
Значение доминантного очага возбуждения в ЦНС заключается в том, что на его базе формируется конкретная приспособительная деятельность, ориентированная на достижение полезных результатов, необходимых для устранения причин, поддерживающих тот или иной нервный центр в доминантном состоянии. Например, на базе доминантного состояния центра голода реализуется пищедобывательное поведение, на базе доминантного состояния центра жажды запускается поведение, направленное на поиск воды. Успешное завершение данных поведенческих актов в конечном итоге устраняет физиологические причины доминантного состояния центров голода или жажды. Доминантное состояние центров ЦНС обеспечивает автоматизированное выполнение двигательных реакций.
4. Компенсация нарушенных функций после повреждения того или иного центра - также результат проявления пластичности ЦНС. Хорошо известны клинические наблюдения за больными, у которых после кровоизлияний в вещество мозга повреждались центры регуляции мышечного тонуса и акта ходьбы. Тем не менее, со временем отмечалось, что парализованная конечность у больных постепенно начинает вовлекаться в двигательную активность, при этом нормализуется тонус ее мышц. Нарушенная двигательная функция частично, а иногда и полностью восстанавливается за счет большей активности сохранившихся нейронов и вовлечения в эту функцию других - «рассеянных» нейронов в коре большого мозга^с подобными функциями. Этому способствуют регулярные (настойчивые, упорные) пассивные и активные движения.
ТОРМОЖЕНИЕ В ЦНС
Торможение - это активный нервный процесс, результатом которого является прекращение или ослабление возбуждения. Торможение вторично относительно процесса возбуждения, так как всегда возникает как следствие возбуждения.
Торможение в ЦНС открыл И. М. Сеченов (1863). В опыте на таламической лягушке он определял латентное время сгибатель-ного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Было показано, что латентное время рефлекса значительно увеличивается, если на зрительный бугор предварительно положить кристаллик поваренной соли. Открытие И. М. Сеченова послужило толчком для дальнейших исследований торможения в ЦНС, при этом было открыто два механизма торможения: пост- и пресинаптическое.
А. Постсинаптическое торможение возникает на постси-наптических мембранах нейрона в результате гиперполяризационного постсинаптическрго потенциала, уменьшающего возбудимость нейрона, угнетающего его способность реагировать на возбуждающие влияния. По этой причине вызванный гиперполяризационный потенциал был назван тормозным постсинаптиче-ским потенциалом, ТПСП'(см. рис. 5.6). АмплитудаТПСП 1-5 мВ, он способен суммироваться.
Возбудимость клетки от ТПСП (гиперполяризационного постси-наптического потенциала) уменьшается потому, что увеличивается пороговый потенциал (МО, так как Екр (критический уровень деполяризации, КУД) остается на прежнем уровне, а мембранный потенциал (Е ) возрастает. ТПСП возникает под влиянием и аминокисло-
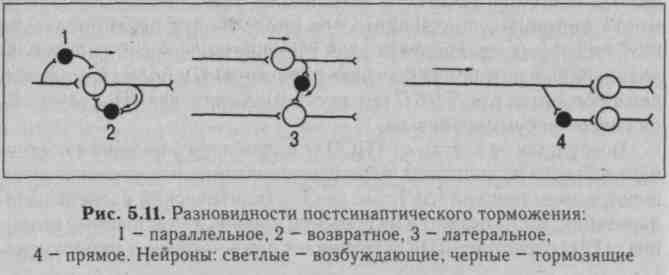
ты глицина, и ГАМК - гамма-аминомасляной кислоты. В спинном мозге глицин выделяется особыми тормозными клетками (клетками Реншоу) в синапсах, образуемых этими клетками на мембране нейрона-мишени. Действуя на ионотропный рецептор постсинапти-ческой мембраны, глицин увеличивает ее проницаемость для СГ, при этом СГ поступает в клетку согласно концентрационному градиенту вопреки электрическому градиенту, в результате чего развивается гиперполяризация. В безхлорной среде тормозная роль глицина не реализуется. Ареактивность нейрона к возбуждающим импульсам является следствием алгебраической суммации ТПСП и ВПСП, в связи с чем в зоне аксонного холмика не происходит деполяризации мембраны до критического уровня. При действии ГАМК на постсинаптическую мембрану ТПСП развивается в результате входа СГ в клетку или выхода К+ из клетки. Концентрационные градиенты ионов К+ в процессе развития торможения нейронов поддерживаются Ыа/К-помпой, ионов СГ - СГ-помпой. Разновидности постсинаптического торможения представлены на рис. 5.11.
|
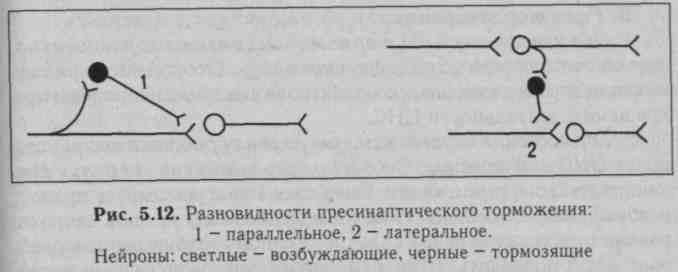
Б. Пресинаптическое торможение развивается в преси-наптических окончаниях. При этом мембранный потенциал и возбудимость исследуемых нейронов не изменяются либо регистрируется низкоамплитудный ВПСП, недостаточный для возникновения ПД (рис. 5.12). Возбуждение блокируется в преси» наптических окончаниях вследствие деполяризации их. В очаге деполяризации нарушается процесс распространения возбуждения, следовательно, поступающие импульсы, не имея возможности пройти зону деполяризации в обычном количестве и обычной амплитуды, не обеспечивают выделение медиатора в синаптическую щель в достаточном количестве, поэтому нейрон не возбуждается, его функциональное состояние, естественно, остается неизменным. Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные клетки, аксоны которых образу-
ют синапсы на пресинаптических окончаниях аксона-мишени (см. рис 5.12). Торможение (деполяризация) после одного афферентного залпа продолжается 300-400 мс, медиатором является гамма-аминомасляная кислота (ГАМК), которая действует на ГАМК-рецепторы.
Деполяризация является следствием повышения проницаемости для СГ, в результате чего он выходит из клетки согласно электрическому градиенту. Это доказывает, что в составе мембран пресинаптических терминалей имеется хлорный насос, обеспечивающий транспорт СГ внутрь клетки вопреки электрическому градиенту.
Разновидности пресинаптического торможения изучены недостаточно. По-видимому, имеются те же варианты, что и для постсинаптического торможения. В частности, на рис. 5.12 представлено параллельное и латеральное пресинаптическое торможение. Однако возвратное пресинаптическое торможение на уровне спинного мозга (по типу возвратного постсинаптического торможения) у млекопитающих обнаружить не удалось, хотя у лягушек
оно выявлено.
В реальной действительности взаимоотношения возбуждающих и тормозных нейронов значительно сложнее, чем представлено на рис. 5.11 и 5.12, тем не менее все варианты пре- и постсинаптического торможений можно объединить в две группы: 1) когда блокируется собственный путь самим распространяющимся возбуждением с помощью вставочных тормозных клеток (параллельное и возвратное торможение) и 2) когда блокируются другие нервные элементы под влиянием импульсов от соседних возбуждающих нейронов с включением тормозных клеток (латеральное и прямое торможения). Поскольку тормозные клетки сами могут быть заторможены другими тормозными нейронами (торможение торможения), это может облегчить распространение возбуждения.
В. Роль торможения.
1.Оба известных вида торможения со всеми их разновидностями выполняют охранительную роль. Отсутствие торможения привело бы к истощению медиаторов в аксонах нейронов и прекращению деятельности ЦНС.
2. Торможение играет важную роль в обработке поступающей в ЦНС информации. Особенно ярко выражена эта роль у пре-синаптического торможения. Оно более точно регулирует процесс возбуждения, поскольку этим торможением могут быть заблокированы отдельные нервные волокна. К одному возбуждающему нейрону могут подходить сотни и тысячи импульсов по разным терми-налям. Вместе с тем, число дошедших до нейрона импульсов определяется пресинаптическим торможением. Торможение латеральных путей обеспечивает выделение существенных сигналов, из фона. Блокада торможения ведет к широкой иррадиации возбуждения и судорогам (например, при выключении пресинаптического торможения бикукулином).
3. Торможение является важным фактором обеспечения координационной деятельности ЦНС.
Дата добавления: 2015-12-22; просмотров: 1162;