ИХ МЕДИАТОРЫ И РЕЦЕПТОРЫ
Мозг человека содержит около 50 миллиардов нервных клеток, взаимодействие между которыми осуществляется посредством множества синапсов, число которых в тысячи раз больше количества самих клеток (1015-1016), так как их аксоны делятся многократно дихотомически, поэтому один нейрон может образовывать до тысячи синапсов с другими нейронами. Нейроны оказывают свое влияние на органы и ткани также посредством синапсов.
А. Нервная клетка (нейрон) является структурной и функциональной единицей ЦНС, она состоит из сомы (тела клетки с яд-
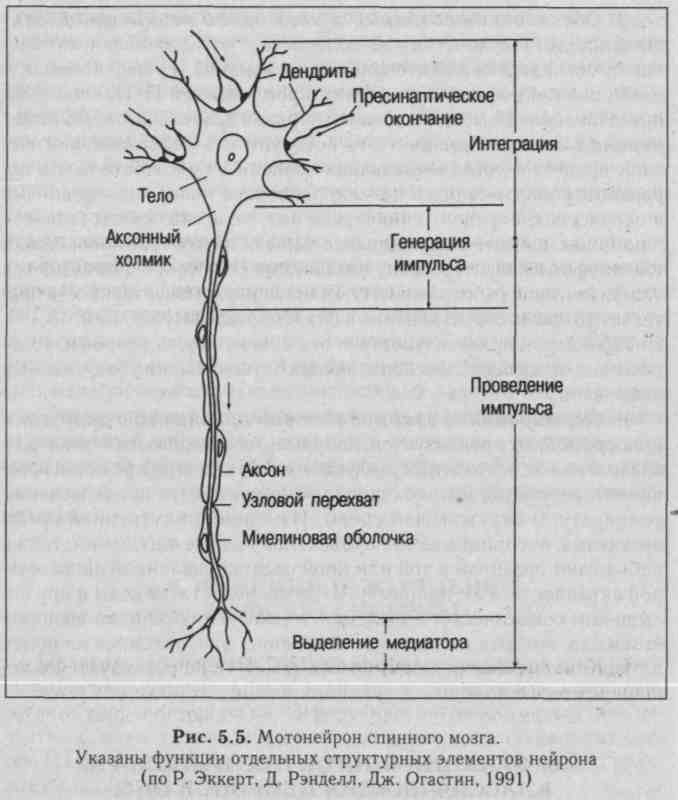
ром) и отростков, представляющих собой большое число дендри-тов и один аксон (рис. 5.5). Потенциал покоя (ПП) нейрона составляет 60-80 мВ, потенциал действия (ПД) -80-110 мВ. Сома и дендриты покрыты нервными окончаниями - синаптическими бутонами и отростками глиальных клеток. На одном нейроне число синаптических бутонов может достигать 10 тысяч (см. рис. 5.5). Аксон начинается от тела клетки аксонным холмиком. Диаметр тела клетки составляет 10-100 мкм, аксона - 1-6 мкм, на периферии длина аксона может достигать метра и более. Нейроны мозга образуют колонки, ядра и слои, выполняющие определенные функции.
Клеточные скопления образуют серое вещество мозга. Между клетками проходят немиелинизированные и миелинизированные нервные волокна (дендриты и аксоны нейронов).
Функциями нервной клетки являются получение, переработка и хранение информации, передача сигнала другим нервным клеткам, регуляция деятельности эффекторных клеток различных органов и тканей организма. Целесообразно выделить следующие функциональные структуры нейрона.
1. Структуры, обеспечивающие синтез макромолекул, - это сома (тело нейрона), выполняющая трофическую функцию по отношению к отросткам (аксону и дендритам) и клеткам-эффекторам. Отросток, лишенный связи с телом нейрона, дегенерирует. Макромолекулы транспортируются по аксону и дендритам.
2. Структуры, воспринимающие импульсы от других нервных клеток, - это тело и дендриты нейрона с расположенными на них шипиками, занимающими до 40% поверхности сомы нейрона и ден-дритов. Причем, если шипики не получают импульсацию, они исчезают. Импульсы могут поступать и к окончанию аксона - аксо-аксонные синапсы, например, в случае пресинаптического торможения.
3. Структуры, где обычно возникает потенциал действия (генераторный пункт ПД), - аксонный холмик.
4. Структуры, проводящие возбуждение к другому нейрону или к эффектору, - аксон.
5. Структуры, передающие импульсы на другие клетки, - синапсы.
Б. Классификация нейронов ЦНС. Нейроны делят на следующие основные группы.
1. В зависимости от отдела ЦНС выделяют нейроны соматической и вегетативной нервной системы.
2. По источнику или направлению информации нейроны подразделяют на: а) афферентные, воспринимающие с помощью рецепторов информацию о внешней и внутренней среде организма и передающие ее в вышележащие отделы ЦНС; б) эфферентные, передающие информацию к рабочим органам - эффекторам; нервные клетки, иннервирующие эффекторы, иногда называют эф-фекторными; эффекторные нейроны спинного мозга (мотонейроны) делят на а- иу-мотонейроны; в) вставочные (интернейроны), обеспечивающие взаимодействие между нейронами ЦНС.
3. По медиатору, выделяющемуся в окончаниях аксонов, различают нейроны адренергические, холинергические, серотонинер-гические и т. д.
4. По влиянию - возбуждающие и тормозящие.
В. Глиальные клетки (нейроглия - «нервный клей») более многочисленны, чем нейроны, составляют около 50% объема ЦНС. Они способны к делению в течение всей жизни. Размеры глиальных клеток в 3—4 раза меньше нервных, с возрастом их число увеличивается (число нейронов уменьшается). Тела нейронов, как и их аксоны, окружены глиальными клетками. Глиальные клетки выполняют несколько функций: опорную, защитную, изолирующую, обменную (снабжение нейронов питательными веществами). Микроглиальные клетки способны к фагоцитозу, ритмическому изменению своего объема (период «сокращения» -1,5 мин, «расслабления» - 4 мин). Циклы изменения объема повторяются через каждые 2-20 час. Полагают, что пульсация способствует продвижению аксоплазмы в нейронах и влияет на ток межклеточной жидкости. Мембранный потенциал клеток нейроглии составляет 70-90 мВ, однако ПД они не генерируют, возникают только лишь локальные токи, электротонически распространяющиеся от одной клетки к другой. Процессы возбуждения в нейронах и электрические явления в глиальных клетках, по-видимому, взаимодействуют."
Г. Ликвор - бесцветная прозрачная жидкость, заполняющая мозговые желудочки, Спинномозговой канал и субарахноидальное пространство. Ее происхождение связано с интерстициальной жидкостью мозга, значительная часть ликвора образуется сосудистыми сплетениями желудочков мозга. Непосредственной питательной средой клеток мозга является интерстициальная жидкость, в которую клетки выделяют также и продукты своего обмена. Лик-вор представляет собой совокупность фильтрата плазмы крови и интерстициальной жидкости: она содержит около 90% воды и около 10% сухого остатка (2% - органические, 8% - неорганические вещества).
Д. Медиаторы и рецепторы синапсов ЦНС. Медиаторами синапсов ЦНС являются многие химические вещества, разнородные в структурном отношении (в головном мозге к настоящему времени обнаружено около 30 биологически активных веществ). Вещество, из которого синтезируется медиатор (предшественник медиатора), попадает в нейрон или его окончание из крови или ликвора, в результате биохимических реакций под действием ферментов в нервных окончаниях превращается в соответствующий медиатор и накапливается в синаптических везикулах. По химическому строению медиаторы можно разделить на несколько групп, главными из которых являются амины, аминокислоты, полипептиды. Достаточно широко распространенным медиатором является ацетилхолин.
Согласно принципу Дейла,один нейрон синтезирует и использует один и тот же медиатор или одни и те же медиаторы во всех разветвлениях своего аксона («один нейрон - один медиатор»). Кроме основного медиатора, как выяснилось, в окончаниях аксона могут выделяться и другие - сопутствующие медиаторы (ко-медиаторы), играющие модулирующую роль и более медленно действующие. Однако в спинном мозге установлено два быстродействующих медиатора в одном тормозном нейроне - ГАМК и глицин и даже один тормозной (ГАМК) и один возбуждающий (АТФ). Поэтому принцип Дейла в новой редакции сначала звучал: «Один нейрон - один быстрый медиатор», а затем: «Один нейрон - один быстрый синаптический эффект» (предполагаются и другие варианты).
Эффект действия медиатора зависит в основном от свойств постсинаптической мембраны и вторых посредников. Это явление особенно ярко демонстрируется при сравнении эффектов отдельных медиаторов в ЦНС и в периферических синапсах организма. Ацетилхолин, например, в коре мозга при микроаппликациях на разные нейроны может вызывать возбуждение и торможение, в синапсах сердца - торможение, в синапсах гладкой мускулатуры желудочно-кишечного тракта - возбуждение. Катехоламины стимулируют сердечную деятельность, но тормозят сокращения желудка и кишечника.
5.7. МЕХАНИЗМ ВОЗБУЖДЕНИЯ НЕЙРОНОВ ЦНС
В любых химических синапсах (ЦНС, вегетативных ганглиях, в нервно-мышечном) механизмы передачи сигнала в общих чертах подобны (см. раздел 2.1). Однако в возбуждении нейронов ЦНС имеются характерные особенности, основными из которых являются следующие.
1. Для возбуждения нейрона (возникновения ПД) необходимы поток афферентных импульсов и их взаимодействие. Это объясняется тем, что один пришедший к нейрону импульс вызывает небольшой возбуждающий постсинаптический потенциал (ВПСП, рис. 5.6) - всего 0,05 мВ (миниатюрный ВПСП). Один пузырек содержит до нескольких десятков тысяч молекул медиатора, например ацетилхолина. Если учесть, что пороговый потенциал нейрона 5-10 мВ, ясно, что для возбуждения нейрона требуется множество импульсов.
2. Место возникновения генераторных ВПСП, вызывающих ПД нейрона. Подавляющее большинство нейрональных синапсов находится на дендритах нейрона. Однако наиболее эффективно вызывают возбуждение нейрона синаптические контакты,
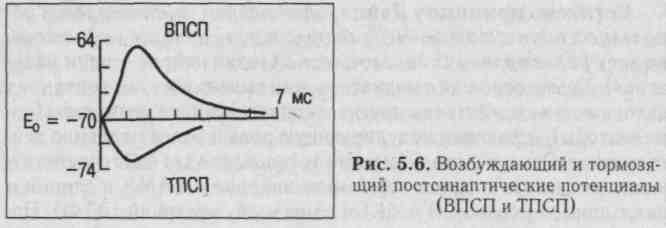
расположенные на теле нейрона. Это связано с тем, что постси-наптические мембраны этих синапсов располагаются в непосредственной близости от места первичного возникновения ПД, располагающегося в аксонном холмике. Близость соматических синапсов к аксонному холмику обеспечивает участие их ВПСП в механизмах генерации ПД. В этой связи некоторые авторы предлагают называть их генераторными синапсами.
3. Генераторный пункт нейрона, т.е. место возникновения ПД, — аксонный холмик. Синапсьг на нем отсутствуют, отличительной особенностью мембраны аксонного холмика является вы-" сокая возбудимость, в 3-4 раза превосходящая возбудимость сома-дендритной мембраны нейрона, что объясняется более высокой концентрацией Ыа-каналов на аксонном холмике. ВПСП элек-тротонически достигают аксонный холмик, обеспечивая здесь уменьшение мембранного потенциала до критического уровня. В этот момент возникает ПД. Возникший в аксонном холмике ПД, с одной стороны, ортодромно переходит на аксон, с другой - антидромно на тело нейрона.
4. Роль дендритов в возникновении возбуждения до сих пор дискутируется. Полагают, что множество ВПСП, возникающих на дендритах, электротонически управляют возбудимостью нейрона. В этой связи дендритные синапсы получили название модуляторных синапсов.
5.8. ХАРАКТЕРИСТИКА РАСПРОСТРАНЕНИЯ ВОЗБУЖДЕНИЯ В ЦНС
Особенности распространения возбуждения в ЦНС объясняются ее нейронным строением - наличием химических синапсов, многократным ветвлением аксонов нейронов, наличием замкнутых нейронных путей. Этими особенностями являются следующие.
1. Одностороннее распространение возбуждения в нейронных цепях, в рефлекторных дугах. Одностороннее распространение возбуждения от аксона одного нейрона к телу или дендритам другого нейрона, но не обратно, объясняется свойствами химических синапсов, которые проводят возбуждение только в одном направлении.
2. Замедленное распространение возбуждения в ЦНС по сравнению с нервным волокном объясняется наличием на путях распространения возбуждения множества химических синапсов. Суммарная задержка передачи возбуждения в нейроне до возникновения ПД достигает величины порядка 2 мс.
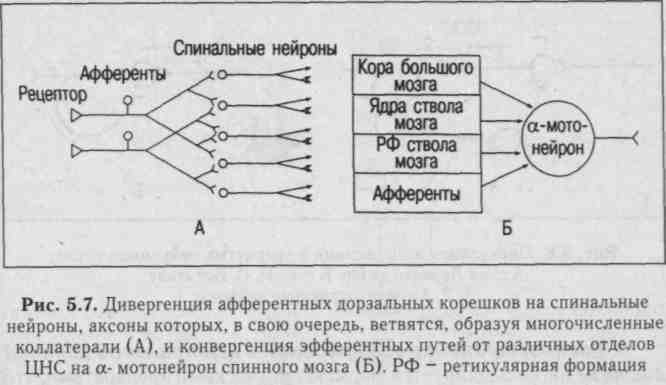
3. Иррадиация (дивергенция) возбуждения в ЦНСобъясняется ветвлением аксонов нейронов, их способностью устанавливать многочисленные связи с другими нейронами, наличием вставочных нейронов, аксоны которых также ветвятся (рис. 5.7 - А).
4. Конвергенция возбуждения (принцип общего конечного пути) - схождение возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному пулу (принцип шеррингтоновской воронки). Объясняется наличием многих аксонных коллатералей, вставочных нейронов, а также тем, что афферентных путей в несколько раз больше, чем эфферентных нейронов. На одном нейроне ЦНС могут располагаться до 10 000 синапсов, на мотонейронах - до 20 000 (рис. 5.7 - Б).
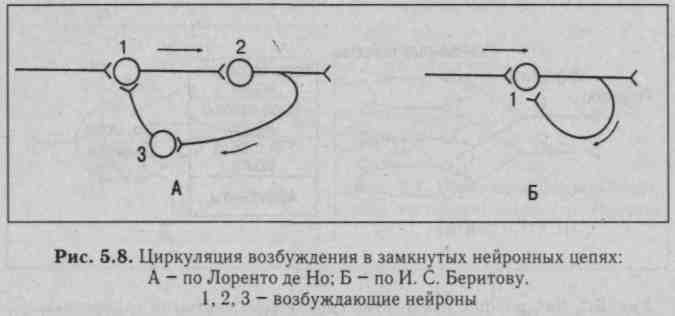
5. Циркуляция возбуждения по замкнутым нейронным цепям, которая может продолжаться минутами и даже часами (рис. 5.8).
6. Распространение возбуждения в центральной нервной системе легко блокируется фармакологическими препаратами, что находит широкое применение в клинической практике. В физиологических условиях ограничения распространения возбуждения по ЦНС связаны с включением нейрофизиологических механизмов торможения нейронов.
Рассмотренные особенности распространения возбуждения дают возможность подойти к пониманию отличительных свойств нервных центров.
Дата добавления: 2015-12-22; просмотров: 1454;