Основные клеточные процессы в онтогенезе. Взаимодействие частей развивающегося зародыша
2.1. Деление клеток
Деление клеток играет большую роль в процессах онтогенеза. Во-первых, благодаря делению из зиготы, которая соответствует одноклеточной стадии развития, возникает многоклеточный организм. Во-вторых, размножение клеток обеспечивает рост организма. В-третьих, избирательному размножению клеток принадлежит заметная роль в обеспечении морфологических процессов. В постэмбриональный период деление клеток осуществляет обновление многих тканей, восстанавливает утраченные органы, заживляет раны.
Зигота, бластомеры и все соматические клетки, за исключением половых делятся митозом. От продолжительности интерфазы митотического цикла зависит частота последовательных делений. В свою очередь интерфаза имеет разную продолжительность в зависимости от стадии развития зародыша, локализации и функции клеток.
Так, в периоде дробления клетки делятся быстрее, чем в другие, более поздние периоды. Во время гаструляции и органогенеза клетки делятся избирательно. В сформированном организме некоторые клетки, например нейроны совсем не делятся, в то время как в кроветворной и эпителиальной тканях продолжается активное размножение клеток, а клетки печени и почек при наличии стимула могут вступить в деление.
В настоящее время известен ряд веществ, которые побуждают клетки к делению, например, фитогемагглютинин, некоторые гормоны, а также комплекс веществ, выделяющихся при повреждении тканей. Открыты также и тканеспецифические ингибиторы клеточного деления – кейлоны. Их действие заключается в подавлении или замедлении скорости деления в тех тканях, которые их вырабатывают.
За последние годы установлено, что многие структуры зародыша образуются клетками, происходящими из небольшого числа или даже одной клетки. Совокупность клеток, являющихся потомками одной родоначальной клетки называют клоном. Например, большие по объему участки центральной нервной системы формируются из определенных клеток раннего зародыша. Остается пока неясным, в какой срок происходит отбор родоначальных клеток и каков механизм этого отбора. Количество циклов клеточных делений в ходе онтогенеза, по-видимому, генетически предопределено.
Таким образом, деление клеток является важным процессом в онтогенезе. Оно протекает с разной интенсивностью в разное время и в разных местах, носит клональный характер и подвержено мутациям. Все это характеризует клеточное деление как сложнейшую функцию целостного организма, подчиняющегося регулирующим влияниям на различных уровнях: генетическом, тканевом, онтогенетическом, биогеоценотическом.
2.2 Миграция клеток
Миграция клеток, или клеточные перемещения, наряду с другими клеточными процессами имеют очень большое значение, начиная с процесса гаструляции и далее, в процессах морфогенеза. Клетки мезенхимного типа мигрируют одиночно и группами, а клетки эпителиев обычно согласованно, пластом. Мезенхима – это скопление веретеновидных или звездчатых клеток, погруженных в межклеточный матрикс. Эпителий – группы клеток, плотно прилежащих друг к другу боковыми стенками и имеющих апикальную и базальную поверхности. Как мезенхима, так и эпителий могут быть образованы из любого из трех зародышевых листков. Клетки мезенхимного типа наиболее подвижны, так как не образуют стойких контактов.
Наиболее яркий пример миграции связан с нервным гребнем. При смыкании нервной трубки клетки нервных валиков выходят из них и располагаются между ее дорзальной частью и эктодермой. Затем они мигрируют в разных направлениях, проявляя очень широкие формообразовательные потенции. Часть клеток нервного гребня в туловищной части зародыша мигрирует в эктодерму и превращается в первичные пигментные клетки - меланоциты. Другие, двигаясь в центральном направлении, образуют нейроны спинальных ганглиев, еще дальше – ганглиев симпатической и парасимпатической систем, третьи превращаются в клетки шванновских оболочек нервов, четвертые – в хромаффинные клетки мозгового вещества надпочечников.
Клетки нервного гребня в головной части зародыша мигрируют в сторону лица, превращаясь в хрящевые, мышечные, соединительнотканные. Они строят хрящи висцерального скелета, мышцы кожи, соединительную ткань лица, языка и нижней челюсти, входят в состав аденогипофиза, паращитовидных желез и мякоти зуба.
Другим убедительным примером выраженных миграций является перемещение первичных половых клеток из желточной энтодермы в зачаток половой железы.
Нарушения миграции клеток в ходе эмбриогенеза приводит к недоразвитию органов или к их гетеротопиям, изменениям нормальной локализации.
Существуют гипотезы, объясняющие движение клеток. Одни клетки движутся по типу хемотаксиса, например гоноциты, сперматозоиды и некоторые клетки крови. Для других клеток более распространенны контактные воздействия. Они представляют собой взаимодействие клеток со структурированным субстратом. Клетки ощущают микроструктуру и движутся вдоль волокон. Хорошо известно, что нервные волокна проходят часто длинный путь по различным тканям от нервного центра к рецептору или эффектору и “узнают” их. Скорее всего нервные окончания растут по микроструктурам субстрата, как они это делают и в культуре тканей.
Таким образом, несомненно, что для миграции конкретных клеток очень важны их способность к амебоидному движению и свойства клеточных мембран. Эти особенности клеток и сама миграция клеток генетически детерминированы с одной стороны, а с другой – находятся под влиянием окружающих клеток и тканей.
2.3. Сортировка клеток
В процессе эмбриогенеза клетки не только активно перемещаются, но и “узнают” друг друга, т.е. создают скопления и пласты только с определенными клетками. Значительные перемещения клеток характерны для периода гаструляции. Смысл этих перемещений заключается в образовании обособленных друг от друга зародышевых листков с совершенно определенным взаимным расположением. Клетки сортируются в зависимости от своих свойств.
Впервые на различия в подвижности и способности к слипанию (адгезии) между клетками, относящимися к разным зародышевым листкам, обратил внимание Г. Гольфретер в 30-х годах этого столетия. Смысл его опытов заключается в том, что зародыши тритонов или других животных на стадии гаструлы диссоциируют с помощью фермента трипсина, который разрушает материал, соединяющий клетки друг с другом. Диссоциированные (дезосегрегированные) клетки тщательно перемешивают, а затем создают такие условия, чтобы клетки могли свободно перемещаться и воссоединяться друг с другом. Сначала клетки представляют собой беспорядочную смесь, затем клетки эктодермы, энтодермы и мезодермы разделяются (сегрегируют) и собираются в отдельные группы, каждая из которых представляет собой клеточный агрегат из однородных клеток. Образуются снова зародышевые листки, располагающиеся иногда в обычном для них порядке.
Подобную сегрегацию клеток можно объяснить способностью к избирательному слипанию клеток одного типа между собой.
Существует несколько гипотез, объясняющих это явление. По одной гипотезе, контакты между однородными клетками сильнее, чем между чужеродными из-за различия в поверхностном заряде их мембран. По другой гипотезе, контактные взаимодействия между одинаковыми клетками основываются на антигенных свойствах их мембран.
По-видимому в сортировке клеток важное место принадлежит генетическому аппарату.
2.4. Гибель клеток
В развитии зародышей наряду с размножением клеток важную роль играют процессы гибели клеток. Примеров, подтверждающих это, достаточно. У головастиков земноводных исчезает хвост, кишечник и жаберные щели, у личинок насекомых разрушаются внутренние органы за небольшим исключением. У человека также имеют место эти процессы. Например, у особей женского пола дегенерируют вольфовы каналы, у особей мужского пола – мюллеровы протоки. Далее, у эмбриона человека вначале закладываются ребра у 7-го шейного позвонка и 9-10 хвостовых позвонков, затем они обычно исчезают, так что шейные позвонки, как правило, ребер не несут и в копчике остается 4-5 позвонков.
Немалое значение принадлежит процессам гибели клеток при образовании полостей тела или сосудов (так называемая кавитация), имеющих в начале вид тяжей без просвета.
В центральной нервной системе сначала образуются больше нервных клеток, чем затем сохраняется, т.к. часть двигательных нейронов, не установивших связь с мышечными волокнами, погибает.
Хорошо изучены процессы гибели клеток при образовании окончательной формы конечностей птиц и млекопитающих. Различные опыты, изучающие гибель клеток позволяют установить, что гибель клеток имеет два уровня регуляции: генетический контроль и межклеточные взаимодействия.
2.5. Дифференцировка клеток
Дифференцировка клеток – это процесс, в результате которого клетка становится специализированной, то есть приобретает химические, морфологические и функциональные особенности. Примером может служить дифференцировка клеток эпидермиса кожи человека, при которой в клетках, перемещающихся из базального в шиповатый и затем в другие, более поверхностные слои, происходит накопление кератогиалина, превращающегося в клетках блестящего слоя в элеидин, а затем в роговом слое – в кератин. При этом изменяется форма клеток, строение клеточных мембран и набор органоидов. Дифференцируется не одна клетка, а группа сходных клеток. В организме человека насчитывается около 100 различных типов клеток. Фибробласты синтезируют коллаген, миобласты – миозин, клетки эпителия пищеварительного тракта пепсин и трипсин и т.д.
Первые химические и морфологические различия между клетками обнаруживаются во время гаструляции. Процесс, в результате которого отдельные ткани в ходе дифференцировки приобретают характерный для них вид, называется гистогенезом. Дифференцировка клеток, гистогенез и органогенез совершаются в совокупности, причем в определенных участках зародыша и в определенное время. Это очень важно, потому что указывает на координированность и интегрированность эмбрионального развития. Возникает вопрос, каким образом клетки, обладающие одинаковым генотипом, дифференцируются и участвуют в гисто- и органогенезе в необходимых местах и в определенные сроки соответственно целостному “образу” данного вида организма. В настоящее время общепризнанной точкой зрения является точка зрения Т. Моргана, который опираясь на хромосомную теорию наследственности, предположил, что дифференцировка клеток в процессе онтогенеза является результатом последовательных реципрокных (взаимных) влияний цитоплазмы и меняющихся продуктов активности ядерных генов. Прозвучала идея о дифференциальной экспрессии генов как основного механизма цитодифференцировки.
В настоящее время собрано много доказательств того, что в большинстве случаев соматические клетки организмов несут полный диплоидный набор хромосом, а генетические потенции ядер соматических клеток также полностью сохраняются, т.е. гены не утрачивают потенциальной функциональной активности. Проведенные цитогенетическим методом исследования кариотипов различных соматических клеток показали почти полную их идентичность. Цитофотометрическим способом установлено, что количество ДНК в них не уменьшается, а методом молекулярной гибридизации показано, что клетки разных тканей идентичны по нуклеотидным последовательностям.
Помимо установленной количественной полноценности ДНК большинства соматических клеток большой интерес представляет вопрос о сохранении функциональных свойств содержащегося в них наследственного материала. Результат многих опытов доказывают это. Так, прошедшая длительный путь дифференцировки соматическая клетка моркови способна развиваться в полноценный организм. У животных отдельные соматические клетки после стадии бластулы, как правило, не способны развиваться в целый нормальный организм, но их ядра, будучи перенесенными в цитоплазму овоцита или яйцеклетки, начинают вести себя соответственно той цитоплазме, в которой они оказались.
Опыты по пересадке ядер соматических клеток в яйцеклетку впервые были осуществлены в 50-х годах в США, а в 60-х годах в Англии английским ученым Д. Гердоном, который используя африканскую шпорцевую лягушку, получил, хотя и в небольшом проценте, но развитие взрослой лягушки из энуклеированной яйцеклетки, в которую пересаживал ядро из эпителиальной клетки кожи лягушки или кишечника головастика. Главный вывод, который вытекал из опыта, заключался в том, что наследственный материал соматических клеток способен сохраняться полноценным не только в количественном, но и в функциональном отношении. Следовательно, цитодифференцировка не является следствием недостаточности наследственного материала. Главная идея заключается в избирательной проявляемости генов в признак, т.е. в дифференциальной экспрессии генов.
Экспрессия гена в признак – сложный этапный процесс, который изучается в основном по продуктам активности гена, с помощью электронного микроскопа или по результатам развития особи.
2.6. Эмбриональная индукция
Эмбриональная индукция – это взаимодействие частей развивающегося зародыша. Главным в эмбриональной индукции является то, что один участок зародыша влияет на судьбу другого участка. Явление эмбриональной индукции с начала ХХ века изучает экспериментальная эмбриология. Классическими считают опыты ученого Г.Шпемена и его сотрудников (1924) на зародышах амфибий, а именно на тритонах, о чем было рассказано выше. Г. Шпемен назвал спинную губу бластопора первичным эмбриональным организатором (индуктором). Первичным потому, что на более ранних стадиях развития зародыша подобных влияний обнаружить не удалось, а организатором потому, что влияние происходило именно на морфогенез. В настоящее время установлено, что главная роль в спинной губе бластопора принадлежит хордомезадермальному зачатку, который назвали эмбриональным индуктором, а само явление, при котором один участок зародыша влияет на судьбы другого – эмбриональной индукцией.
Явление индукции многочисленны и разнообразны. Помимо первичной индукции со стороны спинной губы бластопора описаны индукционные влияния на более поздних, нежели гаструляция, этапах развития. Все они являются вторичными и третичными, представляя собой каскадные взаимодействия, типичные для дифференцировки. Примером вторичной индукции может служить действие глазного бокала на прилежащий покровный эпителий, под влиянием чего эпителий впячивается, а затем отшнуровывается хрусталиковый пузырек – зачаток глазного хрусталика. Расположенный над хрусталиком покровный эпителий тоже испытывает сложные изменения, теряет пигмент и становится роговичным эпителием. Это пример третичной индукции. Таким образом получается, что глазной бокал возникает только после развития передней части головного мозга, хрусталик – после формирования бокала, а роговица – после образования хрусталика, т.е. индукция носит каскадный характер. Но вместе с тем индукция носит и переплетающийся характер, т.е. в индукции той или иной структуры может участвовать не одна, а несколько тканей. Например, глазной бокал служит главным, но не единственным индуктором хрусталика. В свою очередь, сформировавшаяся структура может служить индуктором для несколько других тканей. Чаще всего близлежащие участки зародыша оказывают взаимное влияние друг на друга.
Различают гетерономную и гомономную индукции.
Гетерономная индукция – индукция, при которой один кусочек зародыша индуцирует другой иной орган (хордомезодерма индуцирует появление нервной трубки и всего зародыша в целом).
Гомономная индукция – индукция, при которой индуктор побуждает окружающий материал к развитию в том же направлении, что и он сам (область нефротома, пересаженная другому зародышу, способствует развитию окружающего материала в сторону формирования головной почки; прибавление в культуру фибробластов сердца маленького кусочка хряща влечет за собой процесс образования хряща).
Чтобы воспринять действие индуктора, компетентная (воспринимающая) ткань должна обладать хотя бы минимальной организацией. Одиночные клетки не воспринимают действие индуктора, а чем больше клеток в реагирующей ткани, тем активнее ее реакция. А вот для оказания индуцирующего действия иногда достаточно лишь одной клетки индуктора.
В настоящее время интенсивно ведутся работы по изучению молекулярных и клеточных механизмов индукции.
2.7. Производные зародышевых листков
Из эктодермы развиваются: нервная система, эпидермис кожи, эпителий кожных и молочных желез, роговые образования (чешуя, волосы, перья, ногти), эпителий слюнных желез, хрусталик глаза, слуховой пузырек, периферические чувствительные аппараты, эмаль зубов.
Из энтодермы: хорда, эпителиальная выстилка кишечного тракта и его производных - печени, поджелудочной железы, желудочных и кишечных желез; эпителиальная ткань, выстилающая органы дыхательной системы и частично мочеполовой, а также секретирующие отделы передней и средней доли гипофиза, щитовидной и паращитовидной желез.
Из мезодермы:из наружной (латеральной) части сомитов, т. е. дерматома, образуется соединительная ткань кожи - дерма. Из средней (центральной) части сомитов, т. е. миотома, образуется поперечно-полосатая скелетная мускулатура. Внутренняя (медиальная) часть сомитов, т. е. склеротом, дает начало опорным тканям, сначала хрящевой, а затем костной (в первую очередь тела позвонков) и соединительной ткани, образующей вокруг хорды осевой скелет.
Ножки сомитов (нефрогонатомы) дают начало органам выделения (почечным канальцам) и половым железам.
Клетки, образующие висцеральные и париетальные листки спланхнотома, являются источником эпителиальной выстилки вторичной полости целома. Из спланхнотома также образуется соединительная ткань внутренних органов, кровеносная система, гладкая мускулатура кишечника, дыхательных и мочеполовых путей, скелетная мезенхима, дающая зачатки скелета конечностей.
Глава 3.
Провизорные органы
Провизорные органы - временные специальные внезародышевые органы, обеспечивающие связь зародыша со средой во время эмбрионального развития.
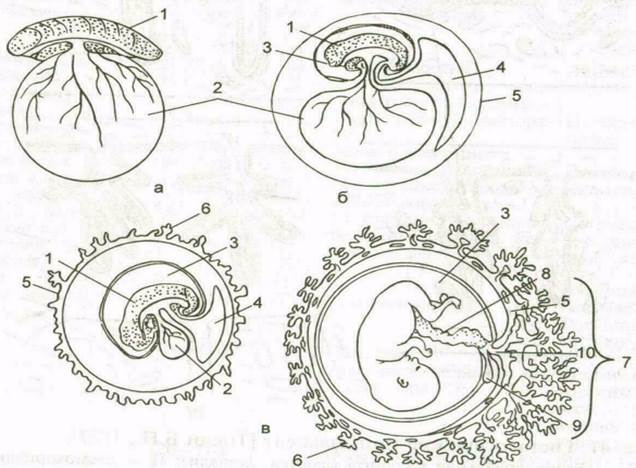
|
Рис. 6. Провизорные органы позвоночных.
А – анамнии; б – неплацентарные амниоты; в – плацентарные амниоты; 1 – зародыш; 2 – желточный мешок; 3 – амнион; 4 – аллантоис; 5 - хорион; 6 – ворсины хориона; 7 – плацента; 8 – пупочный канатик; 9 - редуцированный желточный мешок; 10 – редуцированный аллантоис.
Так как эмбриональное развитие организмов с разным типом развития (личиночным, неличиночным, внутриутробным) протекает в различных условиях, то степень развития и функции провизорных органов у них различны.
3.1. Желточный мешок
Желточный мешок характерен для всех животных с неличиночным типом развития, яйца которых богаты желтком (рыбы, рептилии, птицы). У рыб желточный мешок образуется из клеточного материала трех зародышевых листков, то есть экто-, энто- и мезодермы. У рептилий и птиц внутренний слой желточного мешка имеет энтодермальное, а наружный - мезодермальное происхождение.
У млекопитающих, хотя и нет в яйцеклетках запаса желтка, желточный мешок имеется. Это может быть связано с его важными вторичными функциями. Образуется он из спланхноплевры, которая возникает из образований мезодермального и энтодермального происхождения. Спланхноплевра расщепляется на внутризародышевую и внезародышевую части. Из внезародышевой части формируется желточный мешок.
В стенки желточного мешка врастают кровеносные сосуды, которые образуют густую капиллярную сеть. Клетки стенки желточного мешка выделяют ферменты, расщепляющие питательные вещества желтка, которые поступают в кровеносные капилляры и далее в организм зародыша. Таким образом, желточный мешок выполняет трофическуюфункцию. Желточный мешок является также местом размножения клеток крови, то есть выполняет кроветворнуюфункцию.
У млекопитающих энтодерма желточного мешка служит местом образования первичных половых клеток. Кроме того, желточный мешок млекопитающих заполнен жидкостью, отличающейся высокой концентрацией аминокислот и глюкозы, что указывает на возможность обмена белков в желточном мешке. У разных млекопитающих желточный мешок развит по-разному: у хищников он большой с сильно развитой сетью сосудов. А у приматов сильно сморщивается и исчезает без остатка до родов.
Судьба желточного мешка у разных животных различна. У птиц к концу инкубации остатки желточного мешка находятся внутри зародыша, после чего он быстро рассасывается и исчезает. У млекопитающих редуцированный желточный мешок входит в состав плаценты.
3.2. Амнион с амниотической жидкостью
У пресмыкающихся и птиц запасов желтка в яйцеклетке много, и развитие идет не в воде, а на суше. Поэтому возникает потребность в обеспечении дыхания, выделения, а также защита зародыша от высыхания. В связи с этим у них на ранних стадиях эмбриогенеза начинается формирование такого провизорного органа, как амнион. Наличие или отсутствие амниона лежит в основе деления позвоночных на две группы: амниот и анамний.
Группу анамний составляют более древние позвоночные животные, развивающиеся исключительно в воде и не нуждающиеся в дополнительных водных оболочках. Это такие классы, как Круглоротые, Рыбы и Земноводные. К группе амниот относятся первично наземные позвоночные, у кого эмбриональное развитие протекает на суше в наиболее сложных условиях. Это три класса: Пресмыкающиеся, Птицы и Млекопитающие.
Амнион представляет собой эктодермальный мешок, заполненный амниотической жидкостью и заключающий в себе зародыш. Амнион имеет и мезодермальный слой из внезародышевой соматоплевры, который дает начало гладким мышечным волокнам.
Амнион осуществляет функцию защиты и обмена от высыхания и механических повреждений, создавая для зародыша наиболее благоприятную и естественную водную среду. Амниотическая оболочка специализирована для секреции и поглощения амниотической жидкости, омывающей зародыш. Амниотическая жидкость, в которую погружен зародыш, представляет собой раствор белков, сахаров, минеральных солей, гормонов и мочевины. В процессе развития состав этой среды меняется. Сокращение гладких мышечных волокон, входящих в состав амниона, вызывает пульсацию амниона. Медленные колебательные движения, сообщаемые при этом зародышу, способствуют растущим частям зародыша не мешать друг другу. Амниотическую жидкость, отходящую перед родами, в акушерской практике называют околоплодными водами.
3.3. Хорион (серозная оболочка)
Амниоты кроме амниона имеют и другие провизорные органы. У рептилий, птиц и млекопитающих - это серозная оболочка (хорион).
Хорион - это самая наружная оболочка, прилежащая к скорлупе или материнским тканям, возникает, как и амнион, из эктодермы и соматоплевры.
Хорион служит для обмена между зародышем и окружающей средой.
У яйцекладущих основная его функция - дыхательный газообмен; у млекопитающих он выполняет более обширные функции, участвуя помимо дыхания в питании, выделении, фильтрации и синтезе веществ, в частности, гормонов.
У плацентарных млекопитающих и человека ворсинки хориона врастают в слизистую оболочку матки - специального органа материнского организма. Место наибольшего разветвления ворсинок хориона и наиболее тесного контакта их со слизистой оболочкой матки носит название плаценты или детского места.В зависимости от уровня контакта ворсины и слизистой оболочки матки различают несколько типов плацент.
В плаценте эпителиохориального типа (полуплаценте), характерной для свиньи, лошади, бегемота, верблюда, китообразных, ворсины хориона погружена в складки слизистой оболочки матки без нарушения целостности эпителия, выстилающего ее просвет.
Для жвачных животных типична десмохориальная (соединительнотканнохориальная) плацента. В этом случае целостность эпителиальной выстилки полости матки нарушается. Ворсины хориона погружены в соединительную ткань стенки органа, однако прямой контакт с кровеносными сосудами отсутствует.
В плаценте эндотелиохориального типа ворсины хориона вступают в непосредственный контакт с эндотелием – клетками, ограничивающими просвет сосудов стенки матки.
Гемохориальная плацента приматов и человека обеспечивает наиболее тесную связь зародыша с организмом матери. В этом случае ворсины хориона омываются кровью, изливающейся из сосудов в силу нарушения их стенок в лакуны, которые образуются в области детского места.
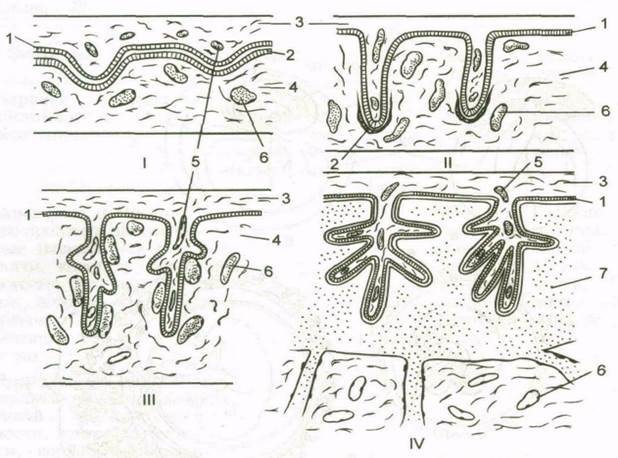
|
Рис. 7. Типы плацент (Токин Б.П., 1987).
I – эпителиохориальная плацента (свиньи, лошади); II - десмохориальная плацента (жвачные); III – эндотелиохориальная плацента (хищники); IV – гемохориальная плацента (насекомоядные, грызуны, приматы
Связь тела зародыша с плацентой осуществляется через пуповину или пупочный канатик, содержащий кровеносные сосуды. Кровеносные капилляры зародыша разветвляются в ворсинках хориона. Так устанавливается плацентарное кровообращение. Кровь матери не смешивается с кровью плода; она омывает ворсинки хориона, не проникает в капилляры плода. Через плаценту плод снабжается питательными веществами, кислородом и освобождается от продуктов жизнедеятельности. Эпителиальные клетки хориона и его ворсинки при этом выполняют важную функцию. Эпителий хориона вместе с клетками стенок сосудов образуют специфический клеточный барьер, через который ряд веществ и микроорганизмы не поступают из кровотока матери в кровоток плода. Нарушение плацентарного барьера может привести к расстройству нормального развития плода. Плацента не является барьером для всех веществ. Некоторые лекарственные вещества, наркотики, пищевые яды, антитела и чужеродные белки могут проходить через плаценту и вызывать патологию беременности. В связи с этим изучение биологических особенностей организма плода и матери у высших млекопитающих и, в первую очередь, у человека имеет большое значение и лежит в основе правильной организации медицинской службы в области охраны материнства и детства.
3.4. Аллантоис.
Аллантоис развивается несколько позже других внезародышевых органов. Он представляет собой мешковидный вырост вентральной стенки задней кишки, то есть он образован энтодермой изнутри и спланхноплеврой снаружи.
У рептилий и птиц аллантоис быстро дорастает до хориона и выполняет несколько функций. Прежде всего он является вместилищем для мочевины и мочевой кислоты, которые представляют собой конечные продукты обмена азотсодержащих органических веществ. В аллантоисе хорошо развита сосудистая сеть, благодаря чему вместе с хорионом он участвует в газообмене. При вылуплении из яйца наружная часть аллантоиса отбрасывается, а внутренняя сохраняется в виде мочевого пузыря.
У многих млекопитающих аллантоис тоже хорошо развит и вместе с хорионом образует хориоаллантоисную плаценту. Термин “плацента” означает тесное слияние зародышевых оболочек с тканями родительского организма.
У приматов энтодермальная часть аллантоиса рудиментирована, а мезодермальные клетки образуют плотный тяж, протягивающийся от клоакального отдела к хориону. По мезодерме аллантоиса к хориону растут сосуды, посредством которых плацента выполняет выделительную, дыхательную и питательную функции. Иначе можно сказать, что аллантоис входит в состав нового специфического провизорного органа - пупочного канатика.
3.5. Критические периоды развития
Критические периоды развития- периоды развития зародыша, наиболее чувствительные к нарушениям условий жизни.
У каждого типа животных наблюдаются свои критические периоды развития. Так, в развитии рыб А.А. Трифонов (1934) и другие исследователи обнаружили три критических периода: первый- начало или середина дробления; второй- начало гаструляции; третий- формирование осевых органов. Развивающиеся икринки в этих стадиях особенно чувствительны к недостатку кислорода, температуре, механическим воздействиям.
Но исследования показывают, что не в любые критические периоды зародыш одинаково чувствителен ко всем повреждающим факторам. Известный эмбриолог П.С. Светлов считает, что в один критический период зародыш более чувствителен к температурным воздействиям, в другой – к химическим. В критические периоды у зародыша сильно изменяется метаболизм, резко усиливается дыхание, меняется содержание РНК, выявляются новые белки, одновременно падает темп роста. Поэтому и является этот период критическим, раз наблюдается такая перестройка в развивающемся зародыше. Всестороннее изучение критических периодов показывает, что они совпадают с активной морфологической дифференцировкой, переходом от одного периода развития к другому, с изменением условия существования зародыша. Есть основания полагать, что разные гены начинают функционировать на различных стадиях онтогенеза, совпадающих с критическими периодами.
У млекопитающих, в частности у человека (по П. С. Светлову, 1959-1962 гг.), следующие критические периоды развития:
1) период имплантации - бластоциста внедряется в стенку матки (6-7 сутки после зачатия) - это период перехода зародыша в новые условия питания и газообмена, требующий новых приспособлений;
2) период плацентации (конец 2-й - начало 3-й недели беременности) –образование третичных ворсин (врастание сосудов в ворсины), период перехода к плацентарному питанию и газообмену;
3) период родов (интранатальный период) - период, когда опять изменяется характер кровообращения, питания и газообмена при переходе из водной среды в воздушную.
Различают критические периоды, общие для всего организма, и периоды в развитии отдельных органах.
Изучение критических периодов в эмбриогенезе показывает необходимость охраны материнского организма от вредных факторов, особенно в первые недели беременности.
Все вредные факторы, вызывающие нарушения в развитии зародыша, называются тератогенными факторами (от греч. terаtos - урод).
Тератогенным действием, как известно, обладают рентгеновские лучи и все другие виды облучения. Если отклоняется температура от нормы или наблюдается кислородное голодание, то также наблюдаются нарушение в развитии зародыша. Тератогенное действие может оказать нехватка веществ, необходимых для нормального метаболизма плода. Так, при острой нехватке витамина А нарушается развитие глаз, сердечно-сосудистой и мочеполовой систем плода. При недостаточности витамина Д нарушается развитие скелета. Прием внутрь хинина, алкоголя, наркотиков, отравления токсическими веществами могут нарушать развитие органов, в первую очередь, нервной системы эмбриона. Иногда после воздействия названных факторов развивается микроцефалия (греч. mikros-малый; kephale-голова); иногда у зародыша полностью отсутствует головной мозг. Подобные нарушения получены экспериментально у животных, подвергшихся аналогичным воздействиям. Тератогенное действие оказывают возбудители некоторых инфекционных и инвазионных болезней. Заражение беременной женщины одноклеточным организмом, токсоплазмой, приводит к нарушению развития плода, чаще всего страдает головной мозг (гидроцефалия, микроцефалия, отставание в умственном развитии вплоть до идиотии). Заболевание беременной женщины вирусом краснухи не отражается существенно на здоровье матери, но может повлиять на развитие зародыша, а, следовательно, и на постэмбриональное развитие. У такого ребенка могут возникнуть пороки в строение сердца, органов зрения и слуха, волчья пасть, заячья губа. Это врожденные нарушения, но они похожи на наследственные нарушения (глухонемота, волчья пасть, заячья губа), которые определяются генотипом и передаются из поколения в поколение.
Тератогенными свойствами могут обладать и некоторые лекарственные препараты. В 50-х годах в Западной Европе беременным женщинам часто назначали снотворное средство «Талидомид», который выпускала одна из западногерманских фармацевтических фирм. Это средство у зародышей вызывало нарушение, которое заключалось в отсутствии или недоразвитии конечностей, нарушениях в строении скелета, лица и других органов. Женские половые гормоны – эстрогены (в частности, синтетический препарат – синэстрол) применяемые у беременных женщин с целью подготовки родовых путей, вызывают развитие опухолей влагалища у девочек, рожденных от этих матерей. Спазмолитический препарат «но-шпа» при применении у беременных женщин вызывает гипоплазию легочной ткани у плода. Применение гербицида “оранжевой смеси” американцами во время войны во Вьетнаме с целью уничтожения листьев и демаскировки партизанских центров, а также газа “си-эс” якобы для выкуривания противника из земляных укрытий и убежищ оказало вред не только огромному числу непосредственно пострадавших от химического оружия во время войны. Прибавляются многие жертвы так называемых отдаленных последствий использования химических средств, у которых развиваются опухолевые заболевания, рак печени и крови. Наблюдаются нарушения нормального течения беременности у женщин и снижение жизнеспособности новорожденных. Большинство этих аномалий является результатом нарушений генетического аппарата, которые произошли под влиянием действия химических средств. Особенно губительным оказалось применение “оранжевой смеси”, содержащей примесь высокотоксического и устойчивого в окружающей среде диоксана, который до сих пор обнаруживается в глубинных слоях почвы. Обращает на себя внимание тот факт, что характер хромосомных аббераций (нарушения в структуре хромосом) и других изменений генетического аппарата у людей, подвергшихся воздействию “оранжевой смеси” сходен с хромосомными нарушениями у жителей Хиросимы и Нагасаки, которые перенесли облучение, вызванное взрывом атомной бомбы. Не пройдет также бесследно взрыв на Чернобыльской АЭС для населения близ расположенных городов и деревень.
Отклонения от нормы, вызванные тератогенными факторами, могут быть различной силы. В зависимости от этого выделяют аномалии, пороки и уродства.
Аномалии - это незначительные отклонения от нормы развития, например, рождение близнецов, наличие жаберных щелей на шее, оволосения.
Пороки развития- резкие отклонения, нарушающие функцию органа и целого организма, снижающие жизнеспособность организма. Например, сросшиеся близнецы, синдактилия, полидактилия.
Уродства -резкие отклонения, снижающие как жизнеспособность организма, так и делающие организм нежизнеспособным. Например, рождение сиреновидных и одноглазых уродов, так называемых "сирен" и "циклопов".
3.6. Антимутагены и их роль
Известно, что в организме есть силы, противодействующие мутагенному действию среды. Это наличие повторов нуклеотидных последовательностей от 10 до 106, вырожденность генетического кода и репарация ДНК. Но также известно, что есть пределы этого противодействия. Во-первых, у разных людей разные генотипы и они по-разному отвечают на изменение окружающей среды, а во-вторых, есть более или менее устойчивые генотипы к действию мутагенов.
Исследования последних десятилетий показали, что имеются факторы, которые обладают не только мутагенным, но и антимутагенным эффектом. Антимутагены – химические соединения, понижающие частоту возникновения мутаций (наследственных изменений организма).
В настоящее время идет активное изучение веществ, организмов, обладающих антимутагенными свойствами (Алекперов У.К., 1984). Поэтому наряду с необходимостью предотвращения загрязнения окружающей среды мутагенными, канцерогенными и тератогенными факторами, очевидна актуальность исследования антимутагенов, эффективно снижающих частоту спонтанных и индуцированных мутаций.
Исследования многих авторов (Алекперов Н.К., 1984; Реймерс М.Ф., 1991 и др.) показывают высокую активность аскорбиновой кислоты (витамин С), ретинола (витамин Е) и его провитамина – β-каротина, токоферола (витамин Е), фолиевой кислоты (витамин В4). Антимутагенная активность указанных веществ связывается с их антиокислительными свойствами.
Витамин С ускоряет репликацию ДНК и размножение клеток, следовательно, процесс восстановления повреждений. Есть данные, что аскорбиновая кислота обладает антиканцерогенной активностью относительно нитрита натрия, используемого в пищевой промышленности. Определение аскорбиновой кислоты, проводимое кафедрой на протяжении десяти лет у беременных женщин г.Владивостока, показывает, что ее уровень ниже нормы, особенно у женщин, родивших детей с врожденными пороками развития (Воронина В.Г., Веревкина Л.В., 1996). Высокое содержание аскорбиновой кислоты отмечается в зеленом и красном перцах, черной смородине, петрушке, апельсиновом и лимонном соках, помидорах, огурцах, клюкве, крыжовнике.
Витамин А и его провитамины – ксантофилл и каротин положительно влияют на зрение и находятся в большом количестве в моркови.
Витамин В4 (фолиевая кислота) служит барьером для вирусов, провоцирующих раковые заболевания. Ежедневная небольшая доза витамина значительно сокращает и даже прекращает развитие предракового состояния у женщин, принимающих пероральные контрацептивы.
Антимутагенные свойства витамина Е (α-токоферола) впервые описаны в 70-х годах. Установлено, что антимутагенное действие токоферола универсально для различных мутагенных факторов химической, физической и биологической природы (Алекперов У.К., 1984). Токоферол содержится в растительных продуктах, маслах, семенах и проростках злаковых, облепихе, паслене, семенах шиповника.
В связи с вышеизложенным особое значение представляет использование природных средств, содержащих указанные витамины, а также другие полезные соединения. К ним относятся многие растения, употребляемые в пищу, особенно дикорастущие. Это крапива, подорожник, щавель, лебеда, шиповник, черника, брусника, ежевика, калина, рябина, земляника, малина, смородина, жимолость, растения семейства аралиевых. Из огородных культур антимутагенным действием обладают укроп, петрушка, морковь, свекла, редька, сельдерей, ревень, спарже, салат, зеленый лук (Свиридов Е.Г., Городинская В.В., 1990)
Экспериментально определено, что токсический эффект снижается под действием яблок в 8 раз, зеленого перца – в 10 раз, баклажан – в 7 раз, винограда – в 4 раза. Рекордсменом оказался лопушник большой – более, чем в 20 раз. Доказано антимутагенное действие кукумарии японской и антиоксидантное действие трепанга (Саватеева Л.Ю., Маслова М.П., Володарский М.В., 1981; Лебская В.Ф. с соавт., 2000; Булгаков А.А. с соавт., 2000 ).
Таким образом, правильное питание является одним из путей предотвращения действия токсических факторов среды. Экспертная группа международной комиссии по охране окружающей среды от мутагенов и канцерогенов отмечает достоверное снижение риска у лиц, придерживающихся диеты, богатой хлебными злаками, овощами, фруктами при снижении потребления продуктов, богатых жирами и алкоголем.
Широкое использование перечисленных средств в процессе трехмесячной подготовки к зачатию при планировании здоровых детей могло бы внести определенный вклад в снижение числа врожденных пороков развития.
3.7. Интегрированность онтогенеза
Организм развивается как целостная система в единстве с условиями среды. Как только в результате дробления образуется два первых бластомера, каждый из них становится неразрывной частью, новой биологической системы и его поведения определяется этой системой. Каждая стадия развития организма есть новое состояние целостности интеграции. Интеграция (лат. integratio – восстановление, восполнение) – объединение в целое каких - либо частей.
На любой стадии развития зародыш представляет собой интегрированное целое, а не сумму бластомеров и клеток. Как указывалось выше, факторами, детерминирующими развитие являются генетические факторы, взаимодействие
частей зародыша, гуморальная, нервная регуляция и воздействие внешних по отношению к зародышу факторов. Генетические факторы запрограммированы в ядре. В любой клетке большинство генов репрессированы, и только часть из них работает. Гены можно подразделить на три группы:
a) гены, функционирующие во всех клетках, например, гены, кодирующие структуры, общие для всех клеток, а также ферменты энергетического обмена;
б) гены, функционирующие в тканях одного типа, например, гены в клетках мышечной ткани регулируют синтез миозина, а в клетках нервной ткани нейропротеина;
в) гены, специфичные для каждого типа клеток, обеспечивающие их морфологию и функции, например, в эритробластах, являющихся предшественниками эритроцитов, работают гены синтеза гемоглобина, а в меланоцитах – гены синтеза меланина.
Взаимодействие частей начинается с того, что возникают различные виды клеток за счет неоднородности цитоплазмы в яйцеклетке. От этого зависит первичная дифференцировка.
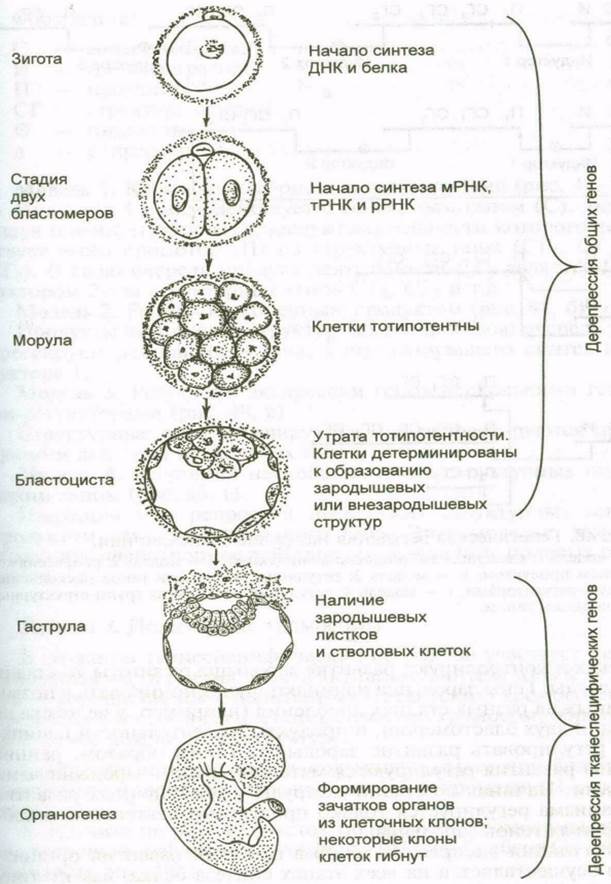
Рис. 8. Генетический контроль развития млекопитающих.
Дальнейшее усложнение в строении частей тела достигается благодаря взаимодействию между клетками.
На определенных стадиях развития клетки зародыша приобретают способность реагировать на влияние других клеток, части развивающегося организма взаимодействуют друг с другом.
Внешними для зародыша факторами, могут быть факторы физической и химической природы, такие как изменения температуры, поступление кислорода, действие лучистой энергии, лекарственных веществ, токсинов, паразитов и др. Различия в уровне интеграции, в характере взаимодействия клеток у разных видов животных могут быть очень существенными. Иногда на молодых стадиях развития зародыш более интегрирован, чем на более поздних стадиях. Так, личинки асцидий, вероятно, более интегрированы, чем взрослые формы. То же, по-видимому, наблюдается у моллюсков и червей. У позвоночных животных интегрированность нарастает по мере углубления процессов органогенеза и цитодифференцировки.
3.8. Особенности эмбрионального развития человека
Яйцеклетки человека содержат небольшое количество желтка. Дробление зиготы у человека полное, но не равномерное. Благодаря асинхронному дроблению могут быть стадии с нечетным числом бластомеров. В результате дробления образуется скопление бластомеров – морула. На стадии 58 бластомеров внутри морулы появляется жидкость, образуется полость (бластоцель), и зародыш превращается в бластоцисту. В бластоцисте, различают наружный слой клеток (трофобласт) и внутреннюю клеточную массу (зародышевый узелок, или эмбриобласт). Внутренняя клеточная масса оттеснена жидкостью к одному из полюсов бластоцисты. Позднее из трофобласта разовьется наружная плодовая оболочка – хорион, а из эмбриобласта – сам зародыш и некоторые внезародышевые органы. Показано, что собственно зародыш развивается из очень небольшого количества клеток зародышевого узелка.
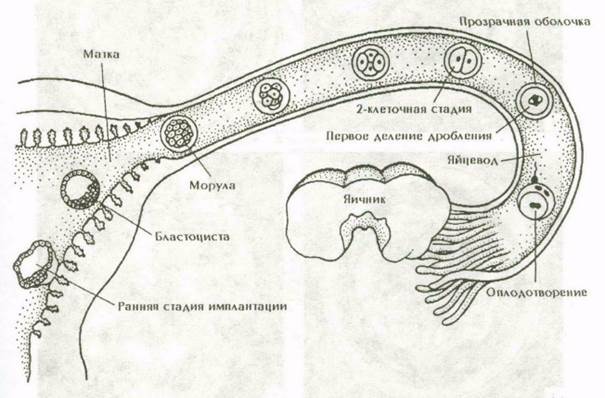
Рис. 9. Овуляция, оплодотворение и человеческий зародыш на первой неделе развития.
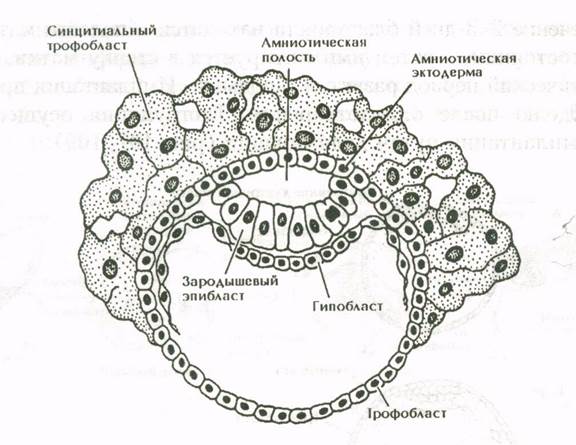
Рис. 10. Бластоциста человека.
Дата добавления: 2015-12-17; просмотров: 9321;