Закономерности проэмбрионального и эмбрионального периодов развития
1.1. Теории онтогенеза
Организмы каждого поколения осуществляют закономерный процесс развития или жизненный цикл. Совокупность взаимосвязанных и детерминированных хронологических событий, закономерно совершающихся в жизненном цикле организма, определяет индивидуальное развитие организма, то есть его онтогенез (греч. ontos – существо, genesis - развитие). Онтогенез начинается от зиготы (при половом размножении) и заканчивается смертью.
В последнее время в онтогенез организма стали включать и образование гамет (мужских и женских), дающих при слиянии зиготу, так как в яйцеклетках находится и-РНК, контролирующая ранние этапы эмбриогенеза.
Единой теории онтогенеза еще не создано, хотя первые сведения о строении зародышей человека и животных были получены учеными Древней Греции.
Знаменитый врач Древней Греции Гиппократ (460 – 370 гг до н. э.) полагал, что в яйце или в теле матери должен находиться маленький, но полностью сформированный организм. Такие убеждения впоследствии стали называться преформизмом (лат. рreformatio-преобразование). В основе преформизма лежали метафизические представления о том, что в онтогенезе никакого развития не существует, а происходит только развертывание и количественное увеличение уже заранее предшествующих частей организма.
Аристотель (384-322 гг. до н. э.), напротив, говорил о том, что организм развивается из бесструктурной гомогенной массы. Это течение получило название эпигенеза (греч. epi.-после, genesis-развитие). Эти взгляды по существу были также метафизическими, так как не учитывалось, что каждое поколение исторически связано с предыдущим, отрицалось преемственность между поколениями.
Эти оба течения - преформизм и эпигенез наибольшего распространения достигли в 17-18 веках.
Нидерландский натуралист Ян Сваммердам (1637-1680) в 1669 г. опубликовал исследования по анатомии насекомых, в котором показал, что куколка содержит в сформированном виде организм бабочки, и сделал предположение о нахождении бабочки также в личинке и яйце. В почках растений Сваммердам обнаружил маленькие настоящие листочки, только в свернутом виде, что также подтверждало его предположение.
Итальянский биолог Марчело Мальпиги (1628-1694), подробно описавший эмбриональное развитие курицы, утверждал, что на всех стадиях развития, начиная от самых ранних, зародыш имеет такое же сложное строение, что и взрослая птица и отличается от нее только размером тела и органов. По его мнению, в яйце с самых ранних стадий развития уже имеется преобразованный зародыш со всеми его органами и процесс развития заключается в количественном изменении, т.е. росте. Развитие зародыша сравнивалось с развитием цветка из бутона, уже содержащего все структурные элементы цветка, только в свернутом, смятом виде.
Наблюдения, проведенные на куколках, насекомых, птицах и почках растений были обобщены и распространены на всех животных и на все растения. Микроскопические исследования яиц и развивающихся зародышей из-за примитивности оптических приборов и несовершенства микроскопической техники не могли опровергнуть положения преформистов. Именно поэтому получили распространения представления о том, что части зародыша, находящиеся в яйце, хотя там и присутствуют, но не различимы, т.к. прозрачны и тесно переплетены.
После того как нидерландский натуралист Антони Левенгук (1632-1723) обнаружил в 1677 г. в семенной жидкости сперматозоиды (анималькули), преформисты разделились на два лагеря. Одни из них овисты (лат. оvum-яйцо) вслед за Мальпиги утверждали, что преобразованный зародыш находится в яйце. Другие – анималькулисты – считали, что яйцеклетка служит только питательным материалом, а преобразованный зародыш находится в сперматозоиде.
Последователь Левенгука голландский микроскопист Николаус Гартсекер (1656-1725) утверждал, что ему удалось увидеть внутри сперматозоида из семенной жидкости мужчины голову, руки, ноги ничтожно маленьких человечков, находящихся под оболочкой головки спермия.
Анималькулисты и овисты по основному вопросу вполне соглашались друг с другом. Изучая процесс развития зародыша, они сводили развитие к чисто количественной стороне – увеличению предшествующих органов, не замечая качественных изменений и отрицая новообразование органов в онтогенезе.
Взгляды преформистов не противоречили религиозной догме творения живых существ Богом. Швейцарский естествоиспытатель и философ Ш. Бонне (1720-1793), стремившийся согласовать данные науки и религии, выступил с теорией вложения. По его представлению, в яичниках “созданной Богом первой женщины Евы” находились все последующие поколения людей, последовательно вложенных друг в друга. И даже некоторые преформисты занимались подсчетом того, сколько поколений было вложено в яйцеклетках мифической прародительницы человечества – Евы.
Первый удар метафизической теории преформизма был нанесен работами академика Российской академии Каспара Вольфа (1733-1794 г.). В своем труде “Теория зарождения” (1759) он описал развитие зародыша цыпленка и доказывал, что на ранних стадиях развития зародыш состоит из однородного студнеобразного вещества, в котором никаких органов нет. Он проследил развитие нервной трубки и пищеварительного канала и показал, что они возникают из первоначально однородной массы. Выступая против преформистов, Вольф развил учения об эпигенезе, согласно которому зародыш испытывает действительное развитие, связанное с возникновением органов заново. Однако учения об эпигенезе не объясняло причины развития зародыша. Почему из однородной массы начинают развиваться органы зародыша? Не находя объяснения этому вопросу, Вольф прибегает к идеалистическому понятию о “существенной силе”, которая, по его мнению, и направляет развитие по определенному пути. Теория Вольфа осталась не понятой его современниками и вскоре была забыта. Лишь в 19 в. она возрождается вновь на более высоком уровне в учении основоположника научной эмбриологии, академика Петербургской академии наук Карла Бэра (1792-1876). Исследуя развитие зародыша курицы, он подтверждает основной вывод Вольфа, что в процессе развития происходит возникновение органов и их усложнение. Однако он не разделял представления Вольфа о чистом эпигенезе.
К. Бэр доказал, что содержимое яйцеклетки неоднородно: оно имеет определенную структуру. По мере развития структурные различия частей зародыша все более увеличиваются. Развитие органов идет путем обособления участков зародышевой ткани с постепенным усложнением и специализацией первоначально близких по строению структур.
Современное понимание развития зародыша отвергает взгляды как преформистов, так и эпигенетиков. Преформизм вообще несовместим с понятием развития. Эпигенез с его представлениями об однородном веществе зиготы и отрицанием в ней каких бы то ни было элементов предопределяющих путь развития, тоже не соответствует фактам. Исследования доказывают, что уже на стадии оплодотворенного яйца конечный результат развития – строение взрослого организма – в значительной мере предопределен, детерминирован (лат. determinare-обусловливать). Из яйца курицы, в каких бы условиях, оно не развивалось (если в этих условиях развития возможно), формируется цыпленок; из зиготы человека развивается человек. Как доказывает генетика, строго определенный путь развития яйца детерминирован наследственными факторами – совокупностью генов, содержащихся в ядре зиготы. Но гены не могут рассматриваться как зачатки органов и тканей. Гены – это участки молекулы ДНК, и ни в структурном, ни в биохимическом отношении не являются копиями детерминированных ими признаков. Однако развитие особи совершается не в пустоте. С самых ранних стадий развития новый организм находится в окружении внешней по отношению к нему среде, с которой он связан. Следовательно, с самого начала развития деятельность генов в клетках находится в тесной зависимости от условий среды.
Для ядер бластомеров средой является цитоплазма, для бластомеров – соседние бластомеры, для закладок органов – организм зародыша, для зародыша (при внутриутробном развитии) – организм матери.
Онтогенез представляет собой последовательное развитие, при котором раннее образовавшиеся структуры обусловливают развитие последующих, причем и эта тенденция проявляется в единстве с условиями среды. Реализация наследственных возможностей организма протекает в определенных условиях и при изменении их происходит по-иному. Однако, совершенно очевидно, что в фенотипе не может быть ни одного признака, который бы не был обусловлен генотипом.
Таким образом, очевидным является то, что именно противоречивое единство внутренних и внешних факторов определяет развитие организма. Так в настоящее время решается вопрос взаимозависимости и взаимообусловленности внешнего и внутреннего в индивидуальном развитии.
1.2. Типы и формыонтогенеза
Существует 2 основных типа онтогенеза: прямой и непрямой (схема 1)
Онтогенез
å æ
Типы: прямой непрямой
å æ â
Формы: неличиночный внутриутробный личиночный
Схема 1
Прямой тип развития наблюдается в двух формах - неличиночного и внутриуробного, а непрямой - в виде личиночного.
Личиночный тип характеризуется наличием одной или нескольких личиночных стадий в развитии организма. Личинки ведут активный образ жизни, сами добывают себе пищу, имеют ряд провизорных (временных) органов, отсутствующих во взрослом состоянии. Этот тип сопровождается превращением (метаморфозом). Личинки покидают яйцевые оболочки до окончания развития и продолжают его вне яйца.
Примером организмов, имеющих личиночную форму развития, являются насекомые, земноводные, низшие хордовые (оболочники).
Неличиночная форма развития имеет место у рыб, пресмыкающихся, птиц, а также у беспозвоночных, яйцеклетки которых богаты желтком. Для питания, дыхания и выделения у этих зародышей также развиваются провизорные органы.
Внутриутробное развитие характерно для высших млекопитающих и человека. Яйцеклетки почти не содержат желтка, все функции зародыша осуществляются через материнский организм. В связи с этим из тканей матери и зародыша образуются сложные провизорные органы, в первую очередь плацента.
1.3. Периоды онтогенеза
В онтогенезе различают 3 периода: проэмбриональный, эмбриональный и постэмбриональный.Для высших животных и человека принято деление на пренатальный (до рождения), интранатальный (рождение) и постнатальный (после рождения) периоды развития (схема 2).
Онтогенез человека
Пренатальный (антенатальный) Интранатальный Постнатальный
(рождение)
     ранний поздний
плод созревание
плода
фетогенез
Эмбриогенез ранний поздний
Бластогенез фетальный фетальный ранний поздний
плод созревание
плода
фетогенез
Эмбриогенез ранний поздний
Бластогенез фетальный фетальный
| Неонатальный ранний | поздний | грудной |
15 дн. 75 дн. 168 дн. 266-272 дн. 23 ч. 59 мин. 28 дн.
(11-12 нед.) (24-25 нед.) (37-38 нед.) (до 6 дн.) (4 нед.)
плод жизнеспособный
плод
Схема 2
1.4. Проэмбриональный период
Проэмбриональный период, предшествующий образованию зиготы, связан с образованием гамет. Иначе, это гаметогенез (овогенез и сперматогенез).
Процессы, характеризующие овогенез, приводят к образованию гаплоидного набора хромосом и образованию сложных структур в цитоплазме. В яйце накапливается р-РНК и и-РНК, идет накопление желтка.От количества желтка и его распределений зависит тип развития онтогенеза.
В зависимости от количества желтка различают 4 типа яйцеклеток:
1) полилецитальные (много желтка; lecitos - гр. желток);
2) мезолецитальные (среднее количество желтка);
3) олиголецитальные (малое количество желтка);
4) алецитальные (желток почти отсутствует).
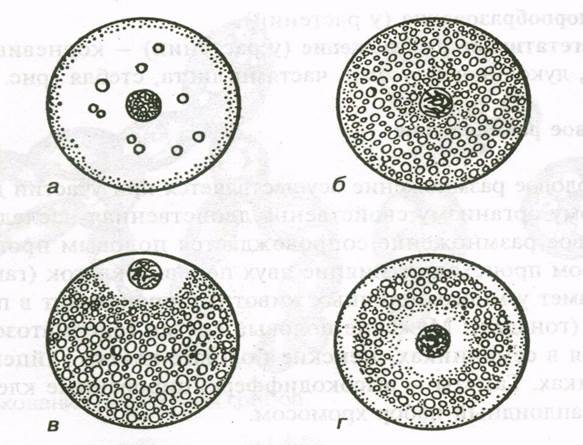
Рис. 1. Типы яйцеклеток по распределению желтка: а – алецитальная, б – изолецитальная, в – телолецитальная, г – центролецитальная.
По характеру распределения желтка яйцеклетки делятся на 3 типа:
1) изолецитальные или гомолецитальные (с равномерным распределением желтка по яйцеклетке), по количеству желтка они чаще олиго- или алецитальные. Примеры: яйца иглокожих, низших хордовых, млекопитающих.
2) телолецитальные (желток сосредоточен на вегетативном полюсе; греч. конец); По содержанию желтка эти яйца чаще всего поли- или мезолецитальные. Примеры: яйца моллюсков, рыб, земноводных, рептилий, птиц.
3) центролецитальные (желток сосредоточен в центре клетки, а цитоплазма по периферии и внутри ядра). Примеры: яйца насекомых, по содержанию желтка это чаще всего олиго- или мезолецитальные яйца.
1.5. Эмбриональный период
Эмбриональный период (греч. еmbryon - зародыш) начинается с оплодотворения и образования зиготы. Окончание этого периода при разных типах онтогенеза связано с различными моментами развития.
Эмбриональный период делится на следующие стадии:
1) оплодотворение – образование зиготы;
2) дробление – образование бластулы;
3) гаструляция – образование зародышевых листков;
4) гисто- и органогенез- образование органов и тканей зародыша.
При личиночной форме онтогенеза эмбриональный период начинается с образования зиготы и заканчивается выходом из яйцевых оболочек.
При неличиночной форме онтогенеза эмбриональный период начинается с образования зиготы и заканчивается выходом из зародышевых оболочек.
При внутриутробной форме онтогенеза эмбриональный период начинается с образования зиготы и длится до рождения.
Зигота.
Зигота- одноклеточная стадия развития нового организма. В зиготе выделяют стадию двух пронуклеусов и стадию синкариона. Стадия двух пронуклеусов предшествует стадии синкариона. Сперматозоид проник в яйцеклетку, но ядра сперматозоида и яйцеклетки еще не слились. Стадия же синкариона характеризуется слиянием ядер. В результате синкариогамии восстанавливается диплоидный набор хромосом. После образования синкариона зигота приступает к дроблению.
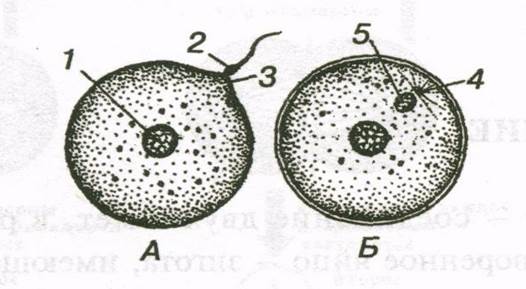
Рис.2. Оплодотворение у млекопитающих. А – сперматозоид проникает в яйцеклетку; Б – из головки сперматозоида образовалось ядро, а из шейки - центриоль. 1- ядро яйцеклетки, 2 – сперматозоид, 3 – воспринимающий бугорок, 4 – центриоль, 5 – ядро сперматозоида.
Дата добавления: 2015-12-17; просмотров: 3172;