Генетические процессы в популяции.
Понятие про популяции. Структура популяций. Роль мутагенного процесса. Закон Харди-Вайнберга. Изменение структуры популяцій.
Одним из важных направлений современной генетики, имеющим большое значение для эволюционного учения, является исследование генетических процессов, протекающих в природных популяциях организмов.
Любой вид живых существ распространен по занимаемому им ареалу не сплошь, а в той или иной мере обособленными друг от друга совокупностями особей — популяциями. Это объясняется прежде всего тем, что условия жизни в разных местах ареала неоднородны и особи данного вида концентрируются преимущественно в наиболее благоприятных для них участках ареала. Таким образом, популяции — это реально существующие ячейки, из которых состоит вид, и именно в них осуществляются начальные этапы эволюционного процесса, ведущие в дальнейшем к возникновению новых видов. Однако не всякая изолированная группа особей представляет популяцию: в пределах популяции особи часто тоже распространены неравномерно, образуя относительно немногочисленные группы, сохраняющиеся лишь в течение короткого времени (на протяжении одного-двух поколений, иногда и того меньше), почему такие группы не обладают собственной эволюционной судьбой, отличной от судьбы остальной части популяции, из которой они временно выделились и с которой быстро снова смешиваются. Наиболее четкое определение популяции можно дать для видов, размножающихся половым путем при перекрестном оплодотворении. У них популяция — это совокупность особей данного вида, в течение длительного времени (большого числа поколений) населяющая определенное пространство, состоящая из особей, могущих свободно скрещиваться друг с другом, и отделенная от таких же соседних совокупностей одной из форм изоляции (пространственной, сезонной, физиологической, генетической).
Закон Харди— Вайнберга. Любая популяция представляет смесь особей, не вполне генотипически одинаковых. Многие гены существуют там в виде различных аллелей и разные особи отличаются друг от друга по этим аллелям. Для понимания генетических процессов, протекающих в популяции, необходимо знать, какие закономерности управляют распределением генов между особями, изменяется ли это распределение из поколения в поколение и если изменятся, то каким образом. Некоторые аспекты этой проблемы были теоретически проанализированы еще в 1908г. одновременно двумя учеными — Г.Харди в Англии и В.Вайнбергом в Германии. Важный итог их работ известен в генетике, как формула Харди — Вайнберга.
Рассмотрим распределение двух аллелей А и а аутосомного гена в популяции перекрестнооплодотворяющихся организмов. Допустим, что численность этой популяции бесконечна велика, популяция полностью изолирована от других популяций этого вида и в ней происходит чисто случайное образование родительских пар, не зависящих от их генотипов и фенотипов (свободное скрещивание или панмиксия). Временно отвлечемся от изменений в составе популяции, могущих возникать вследствие мутаций, превращающих аллель А в аллель а или аллель а валлель А, либо в результате отбора, обусловленного возможными различиями в жизнеспособности или плодовитости особей разного генотипа (АА, Аа и аа).
В такой гипотетической панмиктической популяции любая особь может скрещиваться с любой другой и все они имеют равную вероятность оставить одинаковое в среднем число потомков. Поэтому можно рассматривать всю совокупность половых клеток, образуемых в популяции, как одно целое. Резулътаты соединения мужских и женских половых клеток, происходящего случайно, будут определяться только частотой разных их типов. Каждая половая клетка содержит лишь один аллель данного гена, следовательно, частота этого аллеля в популяции будет равна частоте несущих его половых клеток.
Допустим, что число доминантной и рецессивной аллелей будет одинаковым и равным 0,5. построив решётку Пеннета убедимся, что в F1 соотношение генотипов будет 0,25АА : 0,5Аа : 0,25аа.
| ♂ ♀ | 0,5А | 0,5а |
| 0,5А | 0,25АА | 0,25Аа |
| 0,5а | 0,25Аа | 0,25аа |
Ясно, что в следующем поколении частота образования гамет с доминантной и рецессивной аллелями тоже будет равновероятна, т.е. 0,5А и 0,5а. В таком случае, если свободное скрещивание сохранится, относительная частота разных генотипов не изменится, и такая же картина генетической структуры популяции будет сохраняться и в последующем. Аналогичные закономерности будут наблюдаться и в тех случаях, когда исходные частоты аллелей неодинаковы. Формула Харди-Вайнберга, отражающая частоту генов в популяциях, имеет следующий вид:
(p+q)2=1 или p2+2pq+q2=1,
где p2 – частота гомозиготного потомства по аллели А; 2pq – частота гетерозигот Аа; q2 – частота гомозиготного потомства по аллели а. Аллели А и а присутствуют в популяции с частотами p и q, сумма которых равна 1, т.е. формулу Харди-Вайнберга можно записать и так:
p2АА+2pqАа+q2аа
Формула Харди-Вайнберга даёт возможность рассчитывать относительную частоту генотипов и фенотипов в популяциях, она пригодна и для тех случаев, когда генетические локусы представлены множественными аллелями.
Согласно формуле Харди-Вайнберга, в идеальной популяции, находящейся в равновесии, доли разных генотипов должны неограниченно долго оставаться постоянными. Однако в реальных популяциях эти доли могут изменяться из поколения в поколение вследствие следующих причин: нарушения панмиксии (отсутствия или ограничения свободы скрещиваний), малочисленности популяции, частичного перемешивания разных популяций, возникновения мутаций, наличия отбора (из-за чего различны шансы оставления потомства особями разного генотипа). Теоретический анализ и исследования генетического строения реальных популяций разных организмов показало, что только две первые из перечисленных причин могут приводить к существенному искажению соотношения частот генотипов, ожидаемого в соответствии с формулой Харди—Вайнберга, Поэтому если популяция достаточно велика и в ней наблюдается панмиксия, то распределение генотипов в такой популяции оказывается близким к требуемому этой формулой. И обратно, обнаружение в реальной популяции соотношения частот генотипов, приближающегося к предсказываемому формулой Харди— Вайнберга, служит показателем того, что эта популяция довольно многочисленна и что в ней осуществляются свободные скрещивания. Приведем два примера, иллюстрирующие приложимость формулы Харди—Вайнберга к реально существующим популяциям.
Изменения генетического строения популяций, вызываемые нарушением панмиксии, дрейфом генов и миграциями. Отсутствие или ограничение панмиксии может очень сильно отражаться на генетическом строении популяций. Рассмотрим крайний случай — полное отсутствие панмиксии, наблюдаемое в популяциях строго самооплодотворяющихся организмов, например растений, размножающихся только самоопылением, Представим себе, что на остров, где нет растений изучаемого самоопыляющегося вида, попало одно растение этого вида, имеющее генотип Аа. В результате самоопыления генотипы в F2 распределятся в соотношении 1AA+2Aa+1aa, т.е. в соответствии с формулой Харди — Вайнберга. Однако уже в следующем поколении соотношение частот генотипов будет иным. Если мы обозначим число поколений буквой п, то соотношение частот генотипов в поколении п будет выражаться формулой
(2n-1)AA + 2Aa + (2n-1) aa
В популяции не будет сохраняться равновесие ее генетического состава требуемые формулой Харди — Вайнберга, а, напротив, распределение частот генотипов будет изменяться от поколения к поколению, причем доля в популяции обоих типов гомозигот (АА и аа) будет непрерывно возрастать за счет уменьшения доли гетерозигот Аа.
Если в популяции панмиксия отсутствует не полностью, как в рассмотренном примере, а лишь ограничена в той или иной степени, то распределение частот генотипов будет тем сильнее отличаться от предсказываемого формулой Харди — Вайнберга, чем менее свободно, происходит скрещивание между составляющими ее особями разного генотипа.
Изменения генетического строения популяции могут вызываться и чисто случайными, статистическими причинами. В реальной популяции численность родителей, дающих следующее поколение, всегда не бесконечно велика, как принималось для идеальной популяции, а ограничена, и поэтому распределение частот генотипов среди них может в силу случайного характера выборки оказаться несколько иным, чем во всей популяции в целом. Изменения такого рода тем значительнее, чем меньше эффективная численностьпопуляции, т.е. число особей, фактически участвующих в размножении. Таким образом, при эффективной численности популяции, равной 2, когда в образовании следующего поколения принимают участие всего только две особи – одна женская и одна мужская и при одинаковой частоте обоих аллелей (p=q=0,5), потеря того или другого из этих аллелей происходит с вероятностью 1/16+1/16=1/8 за поколение т.е. с этой вероятностью популяция становится гомозиготной по рассматриваемому гену, другими словами, можно ожидать, что в среднем всего через восемь поколений в популяции сохранится только либо аллель А, либо аллель а. Этот процесс изменения частот генов, в малочисленных популяциях, обязанный случайному сочетанию пар при размножении, получил название дрейфа генов.
Роль дрейфа генов в изменении генетического строения популяции быстро падает с ростом её эффективной численности. Математический анализ этого процесса показывает, что частота гетерозигот в популяции уменьшается за поколение вследствие дрейфа генов на величину, выражаемую формулой 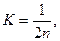 где К – доля, на которую уменьшается частота гетерозигот (или что то же, возрастает частота гомозигот), а п — эффективная численность популяции.
где К – доля, на которую уменьшается частота гетерозигот (или что то же, возрастает частота гомозигот), а п — эффективная численность популяции.
Реальные популяции в пределах вида сравнительно редко бывают полностью изолированными; большей частью происходит некоторое передвижение особей из одной популяции в другую (миграции). Такое передвижение может быть не только активным, но и пассивным (например, перенос семян ветром или птицами). Иногда человек умышленно частично перемешивает популяции. Так, в Сибири, для улучшения местных соболей, в их популяции выпускали баргузинских соболей, обладающих очень темной окраской, ценимой в меховой промышленности. Включение новых групп в исторически сложившиеся популяции постоянно имеет место и в популяциях самого человека. Изменения частот генов, связанные с миграцией особей из популяции в популяцию, называют «перетеканием генов». Изменения эти тем значительнее, чем большую долю составляют иммигранты в популяции, в которую они вливались, и чем сильнее различаются частоты генов в основной популяции и среди иммигрантов.
Влияние мутаций на генетическое строение популяций.Во всякой популяции постоянно возникают мутации различных генов. Хотя спонтанные мутации каждого гена происходят с небольшой частотой, но суммарная роль их в генетике популяций очень велика. Мутационный процесс не только представляет основу разнообразия генов в популяциях, но может существенно влиять на генетическое строение популяции, т.е. на соотношение в ней частот разных генов.
Большинство возникающих в популяции мутаций обречено в силу статистических причин на быстрое исчезновение, но некоторые из них могут сохраняться в течение ряда поколений. Это создает возможность для повышения в популяции частоты тех или иных мутантных генов либо потому, что они будут подхвачены отбором, либо вследствие их случайного дрейфа при колебаниях численности популяции. Кроме того, исчезновению мутантных генов из популяции противостоит действие мутационного процесса, благодаря которому эти же гены появляются там вновь и вновь.
Изменения генетического строения популяций, вызываемые отбором. Единственной силой, направляющей эволюцию органического мира, является естественный отбор. Это направляющее действие отбора проявляется и на начальных этапах эволюционных изменений, протекающих в популяциях и ведущих к внутривидовой дифференциации и образованию новых видов. Поэтому очень важно знать, как влияет отбор на генетическое строение популяций.
Отбор означает дифференциальную вероятность оставления потомства разными особями или группами особей. Вероятность дать потомство детерминируется многими свойствами организма — его жизнеспособностью, быстротой достижения репродуктивного возраста, продолжительностью репродуктивного периода, способностью к скрещиванию, плодовитостью и т.д. Совокупность этих свойств называется приспособленностью особи к условиям среды, в которой они обитают. Как и другие фенотипические характеристики организма, приспособленность в значительной мере определяется генотипом, поэтому генотипически различным особям обычно присуща разная приспособленность. Действие отбора на генетическое строение популяции состоит в том, что некоторые группы особей, генотипически отличающиеся от других, частично ил полностью устраняются от размножения, так что на генетическое строение будущего поколения оказывает влияние только оставшаяся часть популяции, обладающая относительно более высокой приспособленностью.
Ход изменения генетического строения популяции различен в зависимости от того, доминантны или рецессивны элиминируемые отбором гены. Когда отбор ведет к устранению от размножения особей, несущих снижающий приспособленность доминантный ген, то частота этого гена падает относительно быстро, так как он всегда фенотипически проявляется и поэтому в каждом поколении находится под контролем отбора. В популяции, где присутствует такой вредный доминантный ген и его рецессивный аллель, отбор в течение немногих поколений резко снижает долю гомозиготных доминантов и повышает долю гомозиготных рецессивов, доля же гетерозигот снижается постепенно и полная их элиминация происходит значительно позже, чем гомозиготных доминантов. Гораздо медленннее изменяется генетическое строение популяции, когда отбор ведет к устранению от размножения гомозигот по рецессивному гену, делающему их менее приспособленными к окружающим условиям, чем гетерозиготы или гомозиготные доминанты. В этом случае под контроль отбора попадают только особи, гомозиготные по рецессивном гену, а в гетерозиготах этот ген ускользает от отбора. Отбор сперва относительно быстро повышает долю гомозиготных доминантов и уменьшает долю гомозиготных рецессивов, но затем процессы эти сильно замедляются, причем особенно медленно снижается доля гомозиготных рецессивов, так как в каждом поколении они вновь выщепляются в потомстве скрестившихся между собой гетерозигот, доля которых долго остается значительной.
Генетическая гетерогенность природных популяций. Со времени первых работ по генетике природных популяций, проведенных Четвериковым и его сотрудниками в 1920-х годах, различными исследователями накоплен весьма значительный материал о генетическом строении природных популяций ряда организмов.
Многочисленными работами было установлено, что все обследованные природные популяции характеризуются большим генотипическим разнообразием составляющих их особей. Эта генетическая гетерогенность популяций выявляется при изучении наследования встречающихся там фенотипических изменений, генетическим анализом фенотипически нормальных особей на присутствие в них скрытых рецессивных мутаций и цитологическими исследованиями, дающими возможность обнаружить хромосомные перестройки. Работы эти показали, что генетическое строение природных популяций разных организмов обладает некоторыми общими для них типичными чертами.
Самая разительная из этих черт — наличие в природных популяциях множества рецессивных мутаций, скрыто присутствующих в гетерозиготных фенотипически нормальных особях. Насыщенность же популяций рецессивными мутациями объясняется крайней редкостью их фенотипического проявления, вследствие чего они ускользают от действия естественного отбора и некоторое время после возникновения сохраняются там независимо от того, как изменяются признаки организма этими мутациями в гомозиготном состоянии.
В природных популяциях распространены не только рецессивные, но и доминантные мутации. Однако, кроме мономорфных природных популяций, известны и популяции, образуемые особями двух или нескольких четко различных фенотипов. Такой диморфизм или полиморфизм популяций может объясняться модификациями (например, касты у муравьев обязаны разному питанию личинок), а может вызываться генетическими различиями особей. Генетически обусловленный диморфизм или полиморфизм популяции всегда представляет результат естественного отбора, но возникат двояким путём: либо потому, что гетерозиготы по каким-нибудь аллелям имеют больше шансов оставить потомство, чем соответствующие гомозиготы, либо отбор попеременно благоприятствует то одному, то другому аллелю в силу переменчивости условий, в которых обитает популяция.
Гетерозиготное состояние мутаций в популяции приводят к тому, что даже при резко сниженных показателях жизнеспособности и плодовитости рецессивных гомозигот мутантные аллели могут устойчиво сохраняться длительное время. При этом у гетерозигот может наблюдаться более высокая жизнеспособность, чем у доминантных гомозигот. Следствием этого будет существование в популяции нескольких генетически различных форм, что создаёт сбалансированный полиморфизм, т.е. устойчивое воспроизведение нескольких фенотипических классов особей, обусловленное преимуществом гетерозигот.
Генетический полиморфизм, обусловленный тем, что естественный отбор попеременно действует в пользу то одного, то другого аллеля, по-видимому, представляет частое явление в природных популяциях разных организмов. Подробно этот тип полиморфизма изучен у божьих коровок и у хомяков.
Природные популяции разных видов дрозофил характеризуются присутствием в них многих особей, гетерозиготных по инверсиям хромосом. Некоторые из инверсий широко и с большой частотой распространены во многих популяциях. Лабораторными опытами установлено, что гетерозиготы по разным инверсиям различаются по своей приспособленности к повышенной и пониженной температуре и влажности.
Генетический полиморфизм нередок и в природных популяциях растений. В популяциях некоторых видов растений наблюдается генетически обусловленная гетеростилия. Так, в природных популяциях обыкновенного первоцвета всегда имеются особи трех типов: с коротким пестиком и длинными тычинками, с длинным пестиком и короткими тычинками и с пестиками и тычинками одинаковой длины.
Генетическая гетерогенность природных популяций приводит к тому, что средняя приспособленность популяции всегда несколько ниже той, которая характеризовала бы данную популяцию, если бы все составляющие ее особи имели генотип, присущий наиболее приспособленным особям. Этот генетический груз,снижающий приспособленность популяции, слагается из двух основных компонентов: во-первых, постоянно происходящего образования менее приспособленных генотипов в результате расщепления и комбинирования генов при скрещивании генотипически разнородных родителей, и, во-вторых, непрерывного возникновения мутаций, в большинстве своем изменяющих фенотип организма в неблагоприятную сторону. Но для вида в целом генетический груз — это плата за возможность дальнейшего совершенствования. Ведь те же факторы, которые создают генетический груз — комбинации генов и мутации — являются единственным источником не только вредных, но и полезных или потенциально полезных изменений генотипа, представляющих, так сказать, мобилизационный резерв наследственной изменчивости, откуда естественный отбор черпает материал для эволюционных преобразований вида.
| <== предыдущая лекция | | | следующая лекция ==> |
| Дозовые нагрузки при проведении радионуклидной диагностики | | | Сцепленное наследование. Хромосомная теория наследственности. |
Дата добавления: 2015-12-16; просмотров: 6217;