МОДЕЛИ ВНУТРИВИДОВОЙ КОНКУРЕНЦИИ
Рассмотрим простейшую из указанных моделей для вида с дискретными периодами размножения, в которой численность популяции в момент времени t равна N, и изменяется во времени пропорционально величине основной чистой скорости воспроизводства R. Такими видами являются, например, большая часть растений, некоторые виды насекомых, у которых разные поколения четко разнесены во времени. Коэффициент R характеризует количество особей, которое воспроизводится в расчете на одну существующую, а также выживание уже существующих. Данная модель может быть выражена уравнением
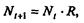 (7.60)
(7.60)
решение которого имеетвид
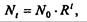 (7.61)
(7.61)
где N0- начальная численность популяции. Эта модель, однако, описывает популяцию, в которой отсутствует конкуренция и в которой R является константой; если R>1, то численность популяции будет бесконечно увеличиваться. В реальности в какой-то момент начинают работать механизмы сдерживания роста популяции. В литературе приводится немало интересных примеров быстрого роста численности популяций, если бы для их размножения существовали идеальные условия. Особенно это относится к насекомым, растениям и микроорганизмам, которые могли бы покрыть земной шар толстым слоем, если им создать благоприятные условия для размножения. Но в действительности такого роста популяций, когда их численность увеличивается в геометрической прогрессии, на сколько-нибудь длительных промежутках времени не наблюдается.
Следовательно, в первую очередь необходимо изменить уравнение (7.60) таким образом, чтобы чистая скорость воспроизводства зависела от внутривидовой конкуренции.
Конкуренцию можно определить как использование некоего ресурса (пищи, воды, света, пространства) каким-либо организмом, который тем самым уменьшает доступность этого ресурса для других организмов. Если конкурирующие организмы принадлежат к одному виду, то взаимоотношения между ними называют внутривидовой конкуренцией, если же они относятся к разным видам, то их взаимоотношения называют межвидовой конкуренцией.
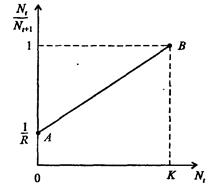
Рис. 7.37. К вопросу об ограничении скорости роста популяции
На рис. 7.37 показана простейшая возможная зависимость скорости воспроизводства от численности популяции. Точка А отражает ситуацию, в которой численность популяции близка к нулю, конкуренция при этом практически отсутствует, и фактическую скорость воспроизводства вполне можно описывать параметром R в его первоначальном виде. Следовательно, при низкой плотности популяции уравнение (7.60) вполне справедливо. В преобразованном виде оно выглядит так:
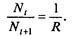
Точка В, напротив, отражает ситуацию, в которой численность популяции высока, и в значительной степени проявляется внутривидовая конкуренция. Фактическая скорость воспроизводства в результате конкуренции настолько снижена, что популяция в целом может не более чем восстанавливать в каждом поколении свою численность, потому что количество родившихся особей уравновешивается количеством погибших. Гипотезе, отраженной на рис. 7.37, соответствует уравнение
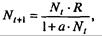 (7.62)
(7.62)
где 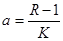 . Это уравнение представляет собой модель роста популяции, ограниченного внутривидовой конкуренцией. Суть этой модели в том, что константа R в уравнении (7.60) заменена на фактическую скорость воспроизводства, т е.
. Это уравнение представляет собой модель роста популяции, ограниченного внутривидовой конкуренцией. Суть этой модели в том, что константа R в уравнении (7.60) заменена на фактическую скорость воспроизводства, т е. 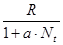 , которая уменьшается по мере роста численности популяции Nt. Достоинство полученного уравнения заключается в его простоте. Такой тип конкуренции приводит к саморегуляции численности популяции, изображенной на рис. 7.38 (для некоторого набора параметров модели; численное решение).
, которая уменьшается по мере роста численности популяции Nt. Достоинство полученного уравнения заключается в его простоте. Такой тип конкуренции приводит к саморегуляции численности популяции, изображенной на рис. 7.38 (для некоторого набора параметров модели; численное решение).
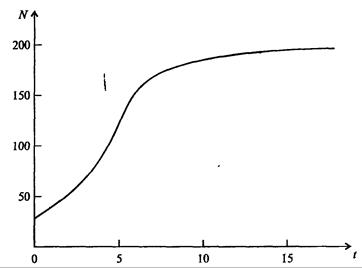
Рис. 7.38. Изменение численности популяции согласно уравнению (7.62) при R = 2, К = 200, N0 = 20
После несложного изменения в уравнении (7.62) может быть получена гораздо более общая модель, учитывающая интенсивность конкуренции. Простейшая из возможных зависимостей падения скорости роста популяции от ее численности, изображенная на рис. 7,37, является не законом природы, а всего лишь удобной гипотезой. Далеко не всегда реальная динамика численности популяции, определяемая внутривидовой конкуренцией, даже качественно согласуется с изображенной на рис. 7.38. Более общая гипотеза о законе падения скорости роста популяции в зависимости от ее численности приводит к следующему уравнению:
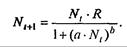 (7.63)
(7.63)
Общность данной модели в отличие от уравнения (7.62) обусловлена введением в модель параметра b, который определяет тип зависимости падения скорости роста популяции от ее численности.
Набор величин a, b, R можно использовать для сравнения и противопоставления сильно различающихся ситуаций. Другим положительным качеством уравнения (7.63) является его способность освещать новые стороны реального мира. Путем анализа кривых динамики популяций, полученных с помощью уравнения, можно прийти к предварительным выводам относительно динамики природных популяций.
На рис. 7.39, а, б, в и г, построенных с помощью численного моделирования, показаны различные варианты динамики численности популяций, полученные с помощью уравнения (7.63) при разном сочетании параметров b и R.
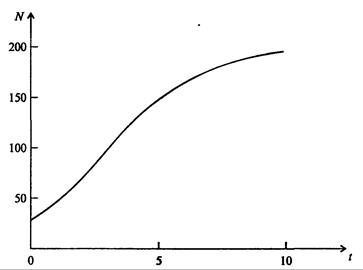
Рис. 7.39, а. Монотонное установление стационарной численности популяции при b =1,4, R =2
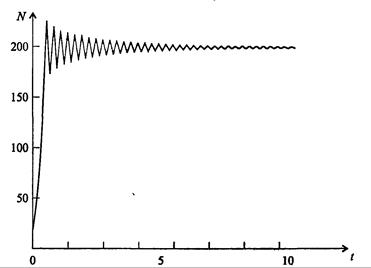
Рис. 7.39, б. Колебательное установление стационарной численности популяции при b =3,9, R =2
Важной частью исследования, связанного с моделью (7.63), является построение на фазовой плоскости (b, R) границ, которые разделяют монотонное затухание, затухающие колебания, устойчивые предельные циклы и случайные (хаотические) изменения, рис. 7.40 Для этого надо задаться значениями а и N0 и производить расчеты, изменяя параметры b, R. Различить каждый из возможных режимов можно попытаться визуально, выполняя построение на экране компьютера графиков изменения численности популяции и запоминая соответствующие значения параметров b, R при переходе от одного режима к другому. Следует, однако, понимать,
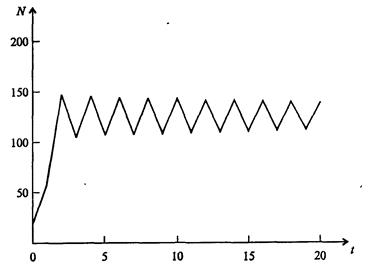
Рис. 7.39, в. Устойчивые предельные циклы изменения численности популяции при b = 3,6, R = 4
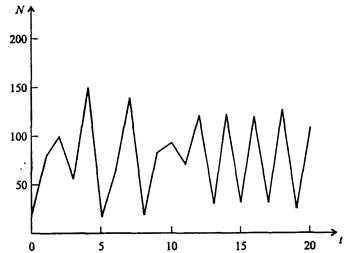
Рис. 7.39, г. Случайные изменения численности популяции при b = 4,4, R = 4
что установление различии между квазипериодическими, апериодическими и хаотическими движениями - сложная математическая задача. Для ее решения используются методы преобразований Фурье и другие, рассмотрение которых выходит за пределы данной книги.
В связи с уравнениями (7.62), (7.63) сделаем замечание общего характера, справедливое для всех моделей в экологии. В отличие от физики, где «модель» часто является точно установленным законом природы, в экологии модель гораздо более условна. Ее адекватность реальности (в той мере, в какой моделирование вообще на это претендует) проверяется лишь экспериментально. Поскольку экологи много раз наблюдали каждую из динамик, изображенных на рис. 7.39, можно сделать вывод о полезности соответствующей модели (иначе она была бы лишь математическим упражнением).
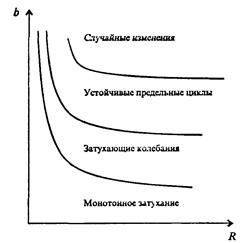
Рис. 7.40. Схематическое изображение фазовой диаграммы динамики численности популяции с дискретным размножением
До сих пор мы рассматривали модели, применимые к популяциям с дискретными периодами размножения. Но в природе встречаются и популяции, где рождение и гибель организмов происходит непрерывно; для таких популяции модели, выраженные уравнениями из предыдущего пункта, непригодны. Рассмотрим популяцию с непрерывным размножением и построим модель изменения ее численности. Математическим аппаратом здесь являются дифференциальные уравнения. Скорость роста в этом случае можно обозначить 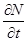 , тогда средняя скорость увеличения численности в расчете на одну особь определяется величиной
, тогда средняя скорость увеличения численности в расчете на одну особь определяется величиной 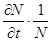 .
.
Без учета внутривидовои конкуренции получаем =rили
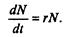
Через r обозначена мгновенная удельная скорость роста численности, т.е. приращение численности за единицу времени в пересчете на одну особь. Согласие этой модели, вследствие того, что не учтена внутривидовая конкуренция, численность популяции при r > 0 будет неограниченно возрастать, т.е. будет наблюдаться экспоненциальный рост.
Теперь попробуем учесть внутривидовую конкуренцию. Для этого воспользуемся методом, который однажды уже применяли (см. рис. 7.37).
Обратимся к рис. 7.41. Когда численность популяции близка к нулю, скоростьростаопределяется величиной r, так как конкуренция еще не оказывает влияния на прирост популяции (точка А). Когда же при возрастании N достигается значение K (предельной плотности насыщения), скорость роста популяции снижается до нуля (точка В).
Записав уравнение прямой АВ, получим
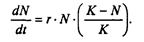 (7.65)
(7.65)
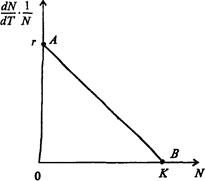
Рис. 7.41. Зависимость скорости воспроизводства от численности популяции
Последнее уравнение известно под названием «логистического». Одно из его достоинств - простота. Уравнение относится к одному из стандартных типов и допускает аналитическое решение:
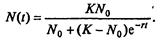 (7.66)
(7.66)
В истории экологии логистическое уравнение сыграло значительную роль, оказав большое влияние на применение в экологии математических методов. С другой стороны, простота уравнения накладывает ограничения на область его применения, так как с его помощью можно описать немногие реальные популяции. Но, несмотря на все ограничения, логистическое уравнение используется как составная часть и в других моделях динамики численности популяции, рассматриваемых ниже.
Изменение численности популяции в соответствии с формулами (7.65)-(7.66) иллюстрирует рис. 7.42.
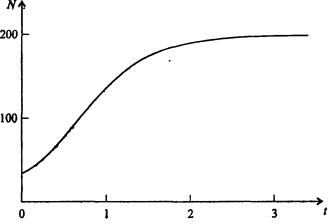
Рис. 7.42. Динамика численности популяции при No =25, К = 200, r = 3
Дата добавления: 2015-10-13; просмотров: 690;