Мозговая основа ощущений
Психологам с 20-х гг. известно, что ощущение возникает довольно поздно – после 100 мс от момента предъявления стимула (т.е. значительно позже прихода сенсорных импульсов в кору). В настоящей работе была поставлена задача понять, что происходит за это время и какой стадии мозговых процессов соответствует возникновение субъективного образа. В исследовании при предъявлении одних и тех же стимулов записывали объективные показатели мозговой деятельности в виде вызванных потенциалов и определяли количественные показатели восприятия. Для получения последних использовались методы теории обнаружения сигнала [Swets et al., 1961], которая описывает процесс восприятия с помощью двух независимых переменных: показателя сенсорной чувствительности d' и критерия решения, определяемого мотивационными факторами. Основные выводы построены на вычислении корреляций между показателями физиологии и психологии, а именно амплитудой волн вызванного потенциала и психофизическими индексами. Исследования проведены на соматосенсорном [Иваницкий, Стрелец, 1976] и на зрительном [Иваницкий, Матвеева, 1976] анализаторах, причем полученные на обеих модальностях результаты были принципиально сходными.
Было установлено, что амплитуда ранних волн вызванного потенциала (ВП) обнаруживала статистически достоверную корреляцию с показателем d', а поздних волн – с критерием решения. Промежуточные волны с латентностью 140 мс для соматосенсорного и 180 мс для зрительного анализатора коррелировали с обоими перцептивными индексами, причем такая двойная корреляция обнаруживалась только для волн проекционной коры. Амплитуда этих волн определялась, таким образом, как сенсорными свойствами стимула, так и его значимостью. На основании имеющихся данных о генезе отдельных волн вызванного потенциала был описан механизм, обеспечивающий указанную двойную корреляцию.
В основе этого механизма лежало кольцевое движение нервных импульсов с «центральной станцией» в проекционной коре. Сначала возбуждение поступало от проекционной к ассоциативной коре. Для зрительных стимулов это были отделы височной коры, играющей важную роль в опознании стимулов, а для кожных раздражений – вторичные и третичные зоны соматосенсорной коры. Затем нервные импульсы приходили к структурам лимбико-гиппокампального комплекса и подкорковым центрам эмоций и мотиваций. До этой стадии поступательное движение возбуждения от рецептивных зон к исполнительным центрам полностью укладывалось в схему рефлекса. Однако процесс на этом не заканчивался: он включал еще возврат возбуждения в кору, включая и ее проекционные отделы (по системе диффузных проекций). Благодаря возврату возбуждения нервные импульсы, приходящие из мотивационных центров, накладывались в нейронах проекционной коры на следы сенсорного возбуждения. На этом этапе или несколько раньше в процесс включалась и лобная кора, что проявлялось в синхронизации рисунка ВП в интервале от 100 до 200 мс после стимула [Иваницкий, Стрелец, 1979]. Было высказано предположение, что указанные промежуточные компоненты ВП отражали синтез на корковых нейронах двух видов информации: наличной информации о физических характеристиках стимула и извлекаемых из памяти сведений о его значимости.
Самое интересное, однако, заключалось в том, что пиковая латентность этих волн ВП точно совпала со временем появления ощущения, измеренным ранее в психологических экспериментах [Froelich, 1929; Pieron, 1961; Бойко, 1964]. Следовательно, можно было предположить, что синтез двух видов информации – наличной и извлекаемой из памяти, – и составляет тот ключевой механизм, который лежит в основе ощущения как феномена уже не физиологического, а психического уровня (рис. 11.1).
Другими словами, был сделан шаг к преодолению барьера между двумя уровнями организации мозговых процессов, один из которых не сопровождается, а другой сопровождается субъективными переживаниями. Согласно предложенной концепции, последовательное поступление информации от рецепторов приводит к повторному движению возбуждения по указанному кольцу, обеспечивая постоянное сопоставление сигналов, приходящих из внешней и внутренней среды, что и составляет психический мониторинг происходящих изменений. Этот процесс осуществляется с периодом квантования приблизительно в 100–180 мс (обратим внимание, что это время сопоставимо с указанным в теории прожектора), составляющим минимальную длительность ощущений [Гольдбурт, Макаров, 1971; Blumenthal, 1977]. Позднее сходное кольцо возбуждения было описано М. Мишкиным [Mishkin, 1993] у обезьян при анализе стимульной информации и образовании памятного следа.
Идея о повторном входе возбуждения в нервные структуры как базисном механизме возникновения субъективного опыта была высказана в последние годы (независимо от нас) рядом авторов. Наиболее полно она была развита в трудах нобелевского лауреата (за исследование антител) Дж. Эделмена [Эделмен, 1981; Edelman, 1989], теория сознания которого получила достаточно широкую известность. По мнению этого автора, в основе возникновения субъективных феноменов лежит механизм повторного входа возбуждения в те же нейронные группы после дополнительной обработки информации в других группах или поступления сигналов из внешней среды, причем обратные связи могут соединять как анатомически близкие, так и удаленные структуры. Этот повторный вход (reentering) дает возможность сравнивать имевшиеся ранее сведения с изменениями, происшедшими в течение одного цикла.
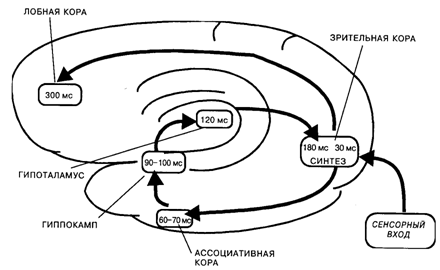
Рис. 11.1. Схема кольцевого движения возбуждения при возникновении зрительных ощущений. Ведущее звено в этом процессе – синтез информации о физических и сигнальных свойствах стимула на нейронах проекционной коры
Кроме того, сходная концепция сознания разрабатывается в нашей стране В. Сергиным [1994], который считает, что данный механизм лежит в основе феномена «внутреннего видения», составляющего суть сознания. Сходные гипотезы о возврате возбуждения в первичную кору как механизме возникновения зрительных и соматосенсорных ощущений были выдвинуты и другими авторами [Cauller, Kulics, 1991; Stoerig, Brandt, 1993; Desmedt, Tomberg, 1995].
Достаточно хорошо проработанная с концептуальной и нейрофизиологической точки зрения концепция сознания была выдвинута Дж. Греем [Gray, 1995]. В соответствии с этой концепцией, содержание сознания определяется активностью субикулярного (отдел гиппокампа) компаратора вместе с обратной связью от данного компаратора к тем наборам нейронов в перцептивной системе, которые снабжают вход данного компаратора с учетом результатов текущего процесса сравнения. Идея о важной роли лимбических структур в генезе психики согласуется и с данными о том, что эти структуры тесно связаны с эксплицитной памятью, узнаванием и вспоминанием [Mishkin et al., 1991]. В то же время вряд ли гиппокампу может принадлежать главная роль в высших функциях мозга. Именно поэтому представления о синтезе информации в коре как ведущем звене в механизмах психики, с нашей точки зрения, более предпочтительны, тем более, что они хорошо согласуются и с данными других авторов, приведенными ранее.
Для оценки предложенных концепций важное значение может иметь соотнесение временной шкалы мозговых процессов со временем субъективно переживаемых событий. По Дж. Эделмену [Edelman, 1989], время одного цикла прохождения возбуждения составляет 100– 150 мс. Если прибавить к этому время, необходимое для прихода сенсорных импульсов в кору, то в сумме получается время, достаточно близкое к тому, которое было найдено в наших экспериментах. Дж. Грей [Gray, 1995] считает, что в квантовании процессов сознания ведущую роль играют процессы, связанные с тета-ритмом, что дает следующее время: 1000/6=166 мс. О значении частот тета-ритма в данном аспекте в свое время писал и П.В. Симонов [1979]. В цитированной ранее работе Дж. Десмедта и К. Томберг [Desmedt, Tomberg, 1995] процесс синхронизации на частоте 40 Гц развивался в течение 100 мс после появления потенциала в первичной коре и до начала волны Р300. Попутно отметим, что синхронизации потенциалов на частоте 40 Гц рядом автором придается большое значение в механизмах сознания и образования зрительных образов [Kulli, Koch, 1991; Engel et al., 1991].
M. Познер и М. Ротбарт [Posner, Rothbart, 1994] показали, что при опознании стимулов вслед за первичной активацией задних отделов коры происходит вовлечение в функцию фронтальной коры с последующим возвратом возбуждения и реактивацией задней коры через 150 мс после ее первоначального возбуждения.
В исследованиях Б. Либета и др. [Libet et al., 1967] с регистрацией вызванных потенциалов на электрокожные стимулы с поверхности коры во время нейрохирургических операций было показано, что на слабые, подпороговые стимулы в коре регистрируются только ранние волны ответа с латентностью до 100 мс. При усилении раздражений в ВП появляются и более поздние колебания с латентностью 150 мс, что сопровождается появлением субъективных ощущений и практически совпадает с латентностью волн ВП, обнаруживших в наших исследованиях двойную корреляцию с перцептивными индексами.
Б. Баарс [Baars, 1993] на основе анализа психологической литературы приходит к выводу, что образы возникают в течение первых 200 мс после действия стимула, а в последующем (200–500 мс) происходит их категоризация.
Из сказанного можно сделать вывод о том, что идея повторного входа возбуждения и информационного синтеза в течение последних 20 лет была высказана независимо друг от друга рядом авторов, относящихся к разным школам и строившим свои заключения на основе разных данных. Очевидно, подобное совпадение не может рассматриваться как случайное. Скорее, оно свидетельствует о приближении к истинному пониманию тех принципов организации мозговых процессов, которые лежат в основе субъективных переживаний.
Приведенные ранее данные касались происхождения одного из наиболее простых психических явлений – ощущений. В связи с этим может возникнуть вопрос о том, насколько данные закономерности универсальны и может ли тот же принцип информационного синтеза быть распространен и на более сложные психические проявления, например на процесс мышления?
Дата добавления: 2015-10-05; просмотров: 564;