Экологическая ниша
Положение вида, которое он занимает в общей системе биоценоза, комплекс его биоценотических связей и требований к абиотическим факторам среды называют экологической нишей вида.
Концепция экологической ниши оказалась очень плодотворной для понимания законов совместной жизни видов. Над ее развитием работали многие экологи: Дж. Гриннелл, Ч. Элтон, Г. Хатчинсон, Ю. Одум и др.
Понятие «экологическая ниша» следует отличать от понятия «местообитание». В последнем случае подразумевается та часть пространства, которая заселена видом и которая обладает необходимыми абиотическими условиями для его существования. Экологическая ниша вида зависит не только от абиотических условий среды, но и в не меньшей мере от его биоценотического окружения. Характер занимаемой экологической ниши определяется как экологическими возможностями вида, так и тем, насколько эти возможности могут быть реализованы в конкретных биоценозах. Это характеристика того образа жизни, который вид может вести в данном сообществе.
Г. Хатчинсон выдвинул понятия фундаментальной и реализованной экологической ниши. Под фундаментальной понимается весь набор условий, при которых вид может успешно существовать и размножаться. В природных биоценозах, однако, виды осваивают далеко не все пригодные для них ресурсы вследствие, прежде всего, конкурентных отношений. Реализованная экологическая ниша – это положение вида в конкретном сообществе, где его ограничивают сложные биоценотические отношения. Иными словами, фундаментальная экологическая ниша характеризует потенциальные возможности вида, а реализованная – ту их часть, которая может осуществиться в данных условиях, при данной доступности ресурса. Таким образом, реализованная ниша всегда меньше, чем фундаментальная.
В экологии широко обсуждается вопрос о том, сколько экологических ниш может вместить биоценоз и сколько видов какой-либо конкретной группы, имеющих близкие требования к среде, могут ужиться вместе.
Специализация вида по питанию, использованию пространства, времени активности и другим условиям характеризуется как сужение его экологической ниши, обратные процессы – как ее расширение. На расширение или сужение экологической ниши вида в сообществе большое влияние оказывают конкуренты. Правило конкурентного исключения, сформулированное Г. Ф. Гаузе для близких по экологии видов, может быть выражено таким образом, что два вида не уживаются в одной экологической нише.
Эксперименты и наблюдения в природе показывают, что во всех случаях, когда виды не могут избежать конкуренции за основные ресурсы, более слабые конкуренты постепенно вытесняются из сообщества. Однако в биоценозах возникает много возможностей хотя бы частичного разграничения экологических ниш близких по экологии видов.
Выход из конкуренции достигается благодаря расхождению требований к среде, изменению образа жизни, что, другими словами, является разграничением экологических ниш видов. В этом случае они приобретают способность сосуществовать в одном биоценозе. Каждый из живущих вместе видов в отсутствие конкурента способен на более полное использование ресурсов. Это явление легко наблюдать в природе. Так, травянистые растения ельника способны довольствоваться небольшим количеством почвенного азота, которое остается от перехвата его корнями деревьев. Однако если на ограниченной площадке обрубить корни этих елей, условия азотного питания трав улучшаются и они бурно идут в рост, принимая густо-зеленую окраску. Улучшение условий жизни и увеличение численности какого-либо вида в результате удаления из биоценоза другого, близкого по экологическим требованиям, называется конкурентным высвобождением.
Разделение совместно живущими видами экологических ниш с частичным их перекрыванием – один из механизмов устойчивости природных биоценозов. Если какой-либо из видов резко снижает свою численность или выпадает из состава сообщества, его роль берут на себя другие. Чем больше видов в составе биоценоза, тем ниже численность каждого из них, тем сильнее выражена их экологическая специализация. В этом случае говорят о «более плотной упаковке экологических ниш в биоценозе».
У близкородственных видов, живущих вместе, обычно наблюдаются очень тонкие разграничения экологических ниш. Так, пасущиеся в африканских саваннах копытные по-разному используют пастбищный корм: зебры обрывают в основном верхушки трав, антилопы гну кормятся тем, что оставляют им зебры, выбирая при этом определенные виды растений, газели выщипывают самые низкие травы, а антилопы топи довольствуются высокими сухими стеблями, оставшимися после других травоядных. Такое же «разделение труда» в южноевропейских степях осуществляли когда-то дикие лошади, сурки и суслики (рис. 92).
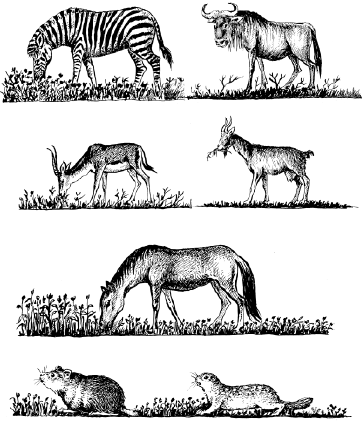
Рис. 92. Разные виды травоядных поедают траву на разной высоте в африканских саваннах (верхние ряды) и в степях Евразии (нижние ряды) (по Ф. Р. Фуэнте, 1972; Б. Д. Абатурову, Г. В. Кузнецову, 1973)
В наших зимних лесах насекомоядные птицы, кормящиеся на деревьях, также избегают конкуренции друг с другом за счет разного характера поиска. Например, поползни и пищухи собирают пищу на стволах. При этом поползни стремительно обследуют дерево, быстро схватывая попадающихся на глаза насекомых или семена, оказавшиеся в крупных трещинах коры, тогда как мелкие пищухи тщательно обшаривают на поверхности ствола малейшие щелки, в которые проникает их тонкий шиловидный клюв. Зимой в смешанных стайках большие синицы ведут широкий поиск на деревьях, в кустах, на пнях, а часто и на снегу; синицы-гаички обследуют преимущественно крупные ветви; длиннохвостые синицы ищут корм на концах ветвей; мелкие корольки тщательно обшаривают верхние части крон хвойных.
Муравьи существуют в природных условиях многовидовыми ассоциациями, члены которых различаются по образу жизни. В лесах Подмосковья чаще всего обнаруживается такая ассоциация видов: доминантный вид (Formica rufa, F. aquilonia или Lasius fuliginosus) занимает несколько ярусов, в почве активен L. flavus, в подстилке леса – Myrmica rubra, напочвенный ярус осваивают L. niger и F. fusca, деревья – Camponotus herculeanus. Специализация к жизни в разных ярусах отражается в жизненной форме видов. Кроме разделения в пространстве, муравьи отличаются и по характеру добывания пищи, по времени суточной активности.
В пустынях наиболее развит комплекс муравьев, собирающих пищу на поверхности почвы (герпетобионтов). Среди них выделяются представители трех трофических групп: 1) дневные зоонекрофаги – активны в самое жаркое время, питаются трупами насекомых и активными днем мелкими живыми насекомыми; 2) ночные зоофаги – охотятся на малоподвижных насекомых с мягкими покровами, которые появляются на поверхности только ночью, и на линяющих членистоногих; 3) карпофаги (дневные и ночные) – поедают семена растений.
Совместно могут обитать по нескольку видов из одной трофической группы. Механизмы выхода из конкуренции и разграничения экологических ниш при этом следующие.
1. Размерная дифференциация (рис. 93). Например, средние веса рабочих особей трех наиболее обычных в песках Кызылкумов дневных зоонекрофагов относятся как 1:8:120. Примерно такое же соотношение весов у некрупной кошки, рыси и тигра.
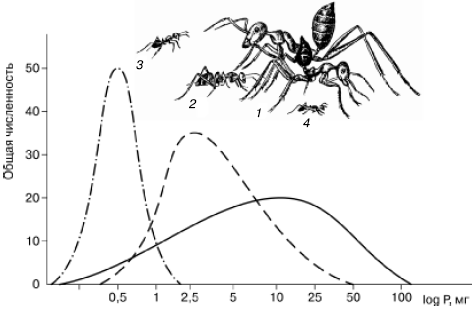
Рис. 93. Сравнительные размеры четырех видов муравьев из группы дневных зоонекрофагов в песчаной пустыне Центральных Каракумов и распределение добычи трех видов по весовым классам (по Г. М. Длусскому, 1981): 1– средний и крупный рабочие Cataglyphis setipes; 2 – С. pallida; 3 – Acantholepis semenovi; 4 – Plagiolepis pallescens
2. Поведенческие различия заключаются в разной стратегии фуражировки. Муравьи, которые создают дороги и используют мобилизацию носильщиков для переноса в гнездо обнаруженной пищи, питаются преимущественно семенами растений, образующих куртины. Муравьи, фуражиры которых работают как одиночные сборщики, собирают в основном семена растений, распределенных дисперсно.
3. Пространственная дифференциация. В пределах одного яруса сбор пищи разными видами может быть приурочен к разным участкам, например на открытых местах или под кустиками полыни, на песчаных или глинистых площадках и т. д.
4. Различия во времени активности относятся преимущественно ко времени суток, но у некоторых видов отмечены несовпадения активности и по сезонам года (преимущественно весенняя или осенняя активность).
Экологические ниши видов изменчивы в пространстве и во времени. Они могут быть резко разграничены в индивидуальном развитии в зависимости от стадии онтогенеза, как, например, у гусениц и имаго чешуекрылых, личинок и жуков майского хруща, головастиков и взрослых лягушек. В этом случае меняется и среда обитания, и все биоценотическое окружение. У других видов экологические ниши, занимаемые молодыми и взрослыми формами, более близки, но тем не менее между ними всегда имеются различия. Так, взрослые окуни и их мальки, живущие в одном и том же озере, используют для своего существования разные энергетические источники и входят в разные цепи питания. Мальки живут за счет мелкого планктона, взрослые – типичные хищники.
Ослабление межвидовой конкуренции приводит к расширению экологической ниши вида. На океанических островах с бедной фауной ряд птиц по сравнению с их сородичами на материке заселяет более разнообразные местообитания и расширяет спектр кормов, так как не сталкивается при этом с конкурирующими видами. У островных обитателей отмечается даже повышенная изменчивость формы клюва как показатель расширения характера кормовых связей.
Если межвидовая конкуренция сужает экологическую нишу вида, не давая проявиться всем его потенциям, то внутривидовая конкуренция, наоборот, способствует расширению экологических ниш. При возросшей численности вида начинается использование дополнительных кормов, освоение новых местообитаний, появление новых биоценотических связей.
Экологические ниши растений, на первый взгляд, менее разнообразны, чем животных. Они четко очерчены у видов, различающихся по питанию: сапрофитов (подъельник, гнездовка), паразитов (заразиха, повилика, раффлезия), полупаразитов (погремок, марьянник, омела), насекомоядных растений (росянка, пузырчатка, непентес). Но и зеленые фотоавтотрофные растения занимают разные экологические ниши в фитоценозах. Множественность экологических ниш определяется разным использованием растениями среды, размещением органов в воздухе и почве, ритмами сезонного развития, длительностью периода вегетации, сроками цветения и особенностями плодоношения, взаимосвязями с элементами абиотической среды и всеми компонентами биоценоза.
В водоемах растения, полностью погруженные в воду (элодея, роголистник, уруть), оказываются в иных условиях температуры, освещенности, газового режима, чем плавающие на поверхности (телорез, водокрас, ряска) или укореняющиеся на дне и выносящие листья на поверхность (кувшинка, кубышка, виктория). Различаются они и взаимосвязями со средой. Эпифиты тропических лесов занимают сходные, но все же не идентичные ниши, так как относятся к разным экологическим группам по отношению к свету и воде (гелиофиты и сциофиты, гигрофиты, мезофиты и ксерофиты). Разные эпифитные орхидеи имеют узкоспециализированных опылителей.
В зрелом широколиственном лесу деревья первого яруса – дуб обыкновенный, вяз гладкий, клен платановидный, липа сердцелистная, ясень обыкновенный имеют сходные жизненные формы. Древесный полог, образованный их кронами, оказывается в одном горизонте, в сходных условиях среды. Но внимательный анализ показывает, что они по-разному участвуют в жизни сообщества и, следовательно, занимают разные экологические ниши. Эти деревья различаются по степени светолюбия и теневыносливости, срокам цветения и плодоношения, способам опыления и распространения плодов, составу консортов и проч. Дуб, вяз и ясень – анемофильные растения, но насыщение среды их пыльцой происходит в разные сроки. Клен и липа – энтомофилы, хорошие медоносы, но цветут в разное время. У дуба – зоохория, у остальных широколиственных деревьев – анемохория. Состав консортов у всех разный.
Если в широколиственном лесу кроны деревьев находятся в одном горизонте, то активные корневые окончания располагаются на разной глубине. Корни дуба проникают наиболее глубоко, выше располагаются корни клена и еще более поверхностно – ясеня. Опад разных видов деревьев утилизируется с разной скоростью. Листья липы, клена, вяза, ясеня к весне почти полностью разлагаются, а листья дуба и весной еще образуют рыхлую лесную подстилку.
В соответствии с представлениями Л. Г. Раменского об экологической индивидуальности видов и с учетом того, что виды растений в сообществе по-разному участвуют в освоении и преобразовании среды и трансформации энергии, можно считать, что в сложившихся фитоценозах каждый вид растения имеет свою экологическую нишу.
В онтогенезе растения, как и многие животные, меняют экологическую нишу. С возрастом они более интенсивно используют и преобразуют среду. Переход растения в генеративный период заметно расширяет круг консортов, меняет размер и напряженность фитогенного поля. Средообразующая роль стареющих, сенильных растений убывает. Они теряют многих консортов, но увеличивается роль связанных с ними деструкторов. Продукционные процессы ослабляются.
У растений имеет место перекрывание экологических ниш. Оно усиливается в отдельные периоды при ограничении ресурсов среды, но поскольку виды используют ресурсы индивидуально, избирательно и с разной интенсивностью, конкуренция в устойчивых фитоценозах ослабляется.
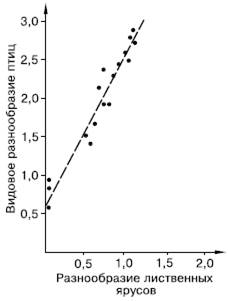
Рис. 94. Корреляция между разнообразием лиственных ярусов и видовым разнообразием птиц (индексы Шеннона по Макартуру из Э. Пианка, 1981)
На богатство экологических ниш в биоценозе оказывают влияние две группы причин. Первая – условия среды, предоставляемые биотопом. Чем мозаичнее и разнообразнее биотоп, тем больше видов могут размежевать в нем свои экологические ниши. Другой источник разнообразия ниш – сами виды, являющиеся ресурсом и создающие среду для других. Американский исследователь Р. Макартур, изучая ярусное разнообразие растительности по горному склону и число видов гнездящихся птиц, продемонстрировал прямую зависимость между этими показателями, положив начало многим исследованиям, подтверждающим связь видовой емкости сообществ с их внутренней структурой (рис. 94). Любой новый вид, внедряющийся в сообщество, увеличивает число имеющихся в нем экологических ниш не только за счет своего собственного положения среди других, но и предоставляя ресурсы для паразитов и хищников, норовых и гнездовых сожителей и т. п.
Дата добавления: 2015-09-23; просмотров: 1232;