Слуховая агнозия и нарушения процесса переработки информации в мозге
Нарушения определения простых и сложных свойств слуховых сигналов.На сонограммах голосов животных, в том числе звуков человеческой речи, представлены три паттерна звуковых частот — постоянные частоты (ПЧ), шум-взрыв (ШВ) и частотно-модулированные (ЧМ) компоненты. Например, сонограмма гласного звука состоит из нескольких горизонтальных линий, или формантов — F1 (самой нижней), F2 и F3. Форманты можно рассматривать как компоненты ПЧ. Сочетание двух фонем, образующее односложное слово, основано на новых компонентах, называемых переходом. Эти сочетания являются ЧМ компонентами. Они очень важны для выделения в словах согласных, которые можно распознать по переходу перед формантами F1 и F2 гласной, например взрывные звуки «к», «т», «п», «г», «д», «б» (Liberman et al., 1956). Вертикальная линия, или полоса частот, показывает разброс интенсивности звука по многим частотам, образующим компоненты шума ШВ, как во фрикативных звуках «с» и «ш». Кроме того, усатые ночницы издают много видов небиосонарных звуков, состоящих из ПЧ, ШВ, или ЧМ компонентов (Kanwal et al., 1993).
Таким образом, звуковые частоты можно считать одним из основных свойств, участвующих в идентификации звука по нейронной репрезентации слуховой информации в слуховом центре. В число других свойств могут входить амплитуда и временная характеристика акустических сигналов, например продолжительность и интервал. Показано, что частотные свойства звуков и их тонотопическая пространственная локализация представлены в первичной слуховой коре летучих мышей (Suga, Jen, 1976), кошек (Merzenith et al., 1975) и обезьян (Brugge, Merzenith, 1973). Определенные нейроны слуховой коры избирательно реагируют на один из трех типов частотных свойств. Они называются ПЧ-, ЧМ- и ШВ-специализированными нейронами (Suga, 1969; Casseday, Covey, 1992). Кроме того, в центральной слуховой системе усатых ночниц была обнаружена диспропорциональная тонотопическая репрезентация при переработке ПЧ-ЧМ сонарных сигналов (Suga & Zen, 1976). Обнаружено, что нейроны, настроенные на опреде-
ленную амплитуду, образуют частотно-амплитудные координаты, общие для всех отделов слуховой коры летучих мышей (Suga, 1977), кошек (Phillips, Orman, 1984) и обезьян (Brugge, Merzenith, 1973). Амплитудно-топическая (ampliotopic) репрезентация обнаружена в отделах слуховой коры у усатых ночниц, кошек (Shreiner et al., 1992) и человека (Pantev et al., 1989).
Простые свойства слуховых сигналов, такие как частота и интенсивность, кодируются периферической слуховой системой. В улитке млекопитающих, в том числе приматов и человека, слуховые клетки имеют тонотопическую организацию вдоль частотной оси базиллярной мембраны. Следовательно, частота акустических сигналов выражается локализацией клеток в улитке, а интенсивность сигналов отражается в скорости разрядов (см. обзор в Suga, 1995). Соответственно, поражения улитки приводят к потере слуха различной степени тяжести. Аналогичные типы потери слуха могут быть вызваны поражениями слуховых ядер в стволе мозга. По-видимому, тонотопическая репрезентация частот и амплиотопическая репрезентация амплитуд не просто фиксирует данные, закодированные периферической слуховой системой. Во многом остается неясным, какую роль играют поражения определенных областей высших уровней слуховой системы в идентификации простых и особенно сложных свойств слуховых сигналов, особенно в слуховой коре человека, поскольку многие описанные результаты были получены в исследованиях усатых ночниц. Кроме того, многие операции, связанные с распознанием свойств слуховых сигналов у человека, невозможно полностью изучить в исследованиях на животных. В их число входят, например, распознание временных паттернов невербальных и особенно вербальных сигналов или переработка сигнала в кратковременной слуховой памяти.
Нейропсихологические исследования процесса центральной переработки слуховой информации у пациентов с локальной патологией мозга дают некоторую дополнительную информацию о нарушениях выделения и распознания некоторых простых и сложных свойств слуховых сигналов, в том числе о нарушениях выделения коротких сигналов, различения простых частот, их временной последовательности и о роли кратковременной рабочей памяти в переработке этой информации.
Нарушения различения слуховых сигналов различной частотности.Роль кратковременной слуховой памяти. В наших работах исследование кратковременной рабочей памяти начиналось с того, что испытуемый учился отличать основной тон в 200 Гц от тона в 300 Гц (Дорофеева, Кайданова, 1969; Тонконогий, 1973; Трауготт, Кайданова, 1975).
Испытуемых просили нажимать кнопку в ответ на тон в 200 Гц и не нажимать кнопку при предъявлении тона в 300 Гц. Выборка пациентов, страдающих последствиями инсульта, состояла из трех групп: 35 пациентов с афазией Вернике и поражением верхнезадней области левой височной доли, 11 пациентов с транскортикальной сенсорной афазией и поражением левой височной доли за пределами области Вернике и 27 пациентов с афазией Брока и поражением премоторной фронтальной области и центральной борозды на уровне третьей фронтальной борозды. У всех пациентов обнаружен нормальный диапазон абсолютных порогов восприятия низко-и среднечастотных тонов. Наблюдаемое возрастание порогов восприятия высоких частот находилось в рамках возрастных норм.
Заучивание сигнала в 200 Гц произошло у пациентов всех групп после 2-3 попыток. Нарушения наблюдались в экспериментальных сериях, где проводилось различение тонов. Обучение различению тонов, различающихся на 100, 200 и 300 Гц, потребовало у пациентов с афазией Вернике 7,6 ± 2,4 попыток, а у пациентов с транскортикальной сенсорной афазией и афазией Брока количество попыток составило 2,7 + 0,8 и 2,7 ± 0,3 соответственно. В следующих сериях основной сигнал в 200 Гц сохранился, но частота сравниваемого сигнала снизилась с 300 до 250, 230, 220, 210 и 205 Гц. Порог различения тонов у пациентов с афазией Вернике достиг 19,2 ± 3,1, а у пациентов с сенсорной афазией и афазией Брока дифференциальные пороги остались на уровне 9-10 Гц и 6-8 Гц соответственно. Увеличение межстимульного интервала от 1 с до 10 с привело к значительному возрастанию дифференциальных порогов у пациентов с афазией Вернике до 38,5 ± 8,2, а у пациентов с сенсорной афазией и афазией Брока пороги слегка возросли до 10-11 Гц.
Интересно, что различению тонов в 200 и 300 Гц в конце концов обучились все 35 пациентов с афазией Вернике, но различению тонов в 200 и 250 Гц, 200 и 230 Гц, 200 и 210 Гц, 200 и 205 Гц обучились соответственно 31,17, 7 и 3 из 35 пациентов с афазией Вернике. Из пациентов с сенсорной афазией отличать тон в 200 Гц от тонов в 300, 250 и 230 Гц научились все 11 испытуемых, а от тона в 210 Гц — 9 из 11 испытуемых. Значительные нарушения были обнаружены только при различении тонов в 200 и 205 Гц. Различать их обучились только 5 пациентов из 11. Все 27 пациентов с афазией Брока обучились различать тона от 300 до 210 Гц. Трудности были обнаружены только при различении стимулов в 200 и 205 Гц у 9 из 27 пациентов. В то же время, различение углов в аналогичной схеме исследования оставалось во всех трех группах пациентов в пределах нормы.
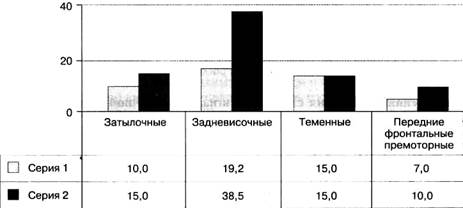
Рис. 2.5.1. Различение тонов пациентами с различной корковой локализацией поражений - затылочной,
задневисочной, теменной и передней фронтальной премоторной: серия 1 - межстимульный интервал
между тонами равен 1 с; серия 2 - интервал увеличен до 10 с
В дальнейших исследованиях различение тонов изучалось у пациентов с затылочными и теменными поражениями (Меерсон, 1986). Получены результаты 10,0 ± 2,8 и 15 ± 3,0 при интервале между тонами l с y пациентов с затылочными
и теменными поражениями соответственно. Эти результаты аналогичны результатам группы пациентов с афазией Брока и сенсорной афазией, и с ними можно сравнить более значительное возрастание порогов от 19,2 ± 3,1 при интервале 1 с до 38,5 ± 8,2 при интервале 10 с у пациентов с афазией Вернике (рис. 2.5.1).
Очевидно, что возрастание порогов различения тонов связано с нарушениями кратковременной слуховой памяти. Нарушения проявляются, когда в ходе обучения требуется удерживать в памяти основной тон в течение всего времени тестирования или на протяжении межстимульного интервала. Пороги значительно возрастают у пациентов с афазией Вернике и верхнезадними височными поражениями в левом полушарии. Как и у пациентов с расстройствами кратковременной зрительной памяти и затылочными поражениями, это возрастание порогов ограничивается достаточно небольшими различиями между тонами по высоте, что указывает на возможное существование двух типов кратковременной слуховой памяти.
Первый тип раскладывает высоту тона на ограниченное число участков с большим разбросом по высоте, достигающим минимум 15-25 Гц. Этот тип памяти лучше защищен от нарушений и более надежен.
Второй тип кратковременной слуховой памяти раскладывает сигнал на большое количество мелких участков, верхняя и нижняя граница которых отстоят друг от друга на 2-3 Гц; они занимают больший объем памяти и более уязвимы к нарушениям. Более точный тип кратковременной слуховой памяти, по-видимому, особенно важен для процесса обучения, при котором для переработки слуховой информации требуется более точное описание простых свойств. Возможно, знакомая слуховая информация обычно перерабатывается с помощью более простого типа кратковременной рабочей памяти.
Различение последовательностей звуков.Распознание слухового потока требует сохранной способности к отбору и группировке последовательностей тонов в потоке, например, различных последовательностей высоких и низких тонов или последовательностей тонов, возрастающих или убывающих по высоте. Различение и распознание таких последовательностей представляет собой часть этого процесса, которая была изучена на пациентах с поражениями мозга.
Эфрон исследовал распознание временной последовательности тонов и щелчков у пациентов с поражениями мозга (Efron, 1963). Согласно наблюдениям автора, нарушение различения последовательностей выявляется только у пациентов с афазией. Исследование Милнер не подтвердило выводы Эфрона (Milner, 1971).
Она предъявляла испытуемым две последовательности из 3 и 5 тонов, полностью идентичные или различающиеся по высоте. Пациент должен был сравнить обе последовательности и сказать, одинаковые они или разные. В группу испытуемых входили пациенты, перенесшие одностороннюю, правую или левую переднемедиальную височную лобэктомию при лечении эпилепсии височной доли. У пациентов, перенесших левостороннюю височную лобэктомию, количество ошибок до и после операции не изменилось. После правосторонней височной лобэктомии среднее количество ошибок увеличилось на 23%. При нарушениях вербальной памяти наблюдается обратная тенденция: количество ошибок возрастает после левосторонней височной лобэктомии.
Однако в исследовании пациентов с афазией Вернике и поражением левого полушария было получено увеличение дифференциальных порогов для второго тона в последовательности из трех тонов (Дорофеева, 1970). Дифференциальный порог у пациентов с афазией Вернике достигал 150 Гц, а у пациентов с афазией Брока составлял только 5-10 Гц. Различия результатов двух исследований могут быть связаны с величиной межстимульных интервалов, которые в эксперименте Милнер были намного короче, благодаря чему последовательности тонов звучали как некоторого рода мелодия. В этом случае задача приближалась к распознанию мелодии, а ее труднее выполнить при поражениях правого полушария. Кроме того, локализация поражений в этих работах различалась. Пациенты, описанные Милнер, перенесли переднемедиальную лобэктомию, а Дорофеева исследовала пациентов после верхнезадних височных инфарктов в левом полушарии, которые привели к афазии Вернике.
В наших исследованиях испытуемым требовалось сравнить последовательности тонов (Тонконогий, 1973; Меерсон, 1986). Последовательности состояли из двух или трех тонов, выстроенных в одинаковом или разном порядке, например 200 Гц — 1000 Гц — 2000 Гц в первой последовательности и 200 Гц — 2000 Гц — 1000 Гц во второй последовательности. Зрительные последовательности состояли из двух или трех незамкнутых контуров, например угол, кривая и прямая. Каждый сигнал в последовательности предъявлялся на 1 с. В первой серии эксперимента интервал между двумя последовательностями составлял 2 с. Во второй и третьей сериях интервал был увеличен до 5 и 10 с. Одинаковые и разные пары последовательностей предъявлялись в случайном порядке. При сравнении последовательностей тонов с интервалом 2 с у пациентов с верхнезадними височными поражениями правого и левого полушария количество ошибок достигало 15,8 и 12,5% соответственно, а при интервале 10 с ответы давались в случайном порядке. При предъявлении зрительных последовательностей со всеми тремя интервалами значительных нарушений не наблюдалось.
У пациентов с лево- и правосторонними затылочными поражениями получены обратные результаты: значительное число ошибок — 14,8 и 10,6% соответственно — для зрительных последовательностей при интервале 2 с и увеличение числа ошибок по мере возрастания интервала до 5 и 10 с. Различение последовательностей тонов было сохранно, число ошибок составляло только 4,8 и 3,2% при лево- и правополушарных поражениях соответственно. У пациентов с поражениями теменной доли зафиксировано значительное число ошибок в обеих последовательностях, зрительной и слуховой: 13,6 и 10,6% для последовательностей контуров и 14,3 и 10,5% для последовательностей тонов при интервале 2 с; увеличение интервала до 5 и 10 с не привело к возрастанию количества ошибок.
Эти данные указывают на важную роль левой и правой височных долей в различении слуховых последовательностей и обеих затылочных долей в различении зрительных последовательностей. По-видимому, наиболее тяжелые поражения височной доли, отражающиеся на различении немелодических слуховых последовательностей, расположены в верхнезадней височной области левого полушария. Относительно недавние исследования показали, что лобэктомия передненижней области правой или левой височной доли приводит к менее серьезным расстройствам кратковременной памяти на слуховые последовательности (Меерсон, 1986). Однако эти расстройства также распространяются на визуальные
последовательности, как и у пациентов с поражениями теменных долей. Это может быть связано с различиями в механизмах нарушений различения последовательностей. Возможно, что у пациентов с верхнезадними височными и затылочными поражениями расстройства различения последовательностей связаны с низшим перцептивным уровнем и определяются трудностями восприятия одиночных свойств последовательностей, высоты тонов в слуховых сигналах и характерных очертаний зрительных сигналов. У пациентов с поражениями передненижних областей височных долей и теменных долей нарушения могут затрагивать высший уровень переработки информации. Высший уровень отвечает за восприятие порядка конкретных сигналов в зрительной или слуховой последовательности как паттерна или специфического «гештальта». Такое симультанное восприятие, возможно, происходит на мультимодальном уровне. Также возможно, что восприятие паттерна или «гештальта» используется на модально-специфических уровнях верхней височной области для звуковых сигналов, а в затылочной доле — для зрительных сигналов.
Роль рабочей памяти в различении последовательностей требует дальнейших исследований. Увеличение межстимульного интервала между двумя сравниваемыми зрительными или слуховыми последовательностями приводит к значительному ухудшению показателей слуховой или зрительной рабочей памяти у пациентов с верхнезадними височными поражениями или затылочными поражениями соответственно.
Конечно, дальнейшие исследования необходимы и для понимания роли, которую играют в развитии невербальной слуховой агнозии описанные расстройства различения слуховых сигналов различной частоты и их последовательностей, особенно в связи с более специфическими типами частот, такими как постоянные частоты (ПЧ), шум-взрыв частоты (noise-burst frequency) (ШВ) и частотно-модулированные компоненты. Связь описанных нарушений со слуховой агнозией вербального типа также будет обсуждаться в разделе 5.1 «Синдромы афазии и другие речевые расстройства».
Нарушения переработки коротких слуховых сигналов.Для выявления элементарного слухового дефицита у пациентов с односторонними корковыми поражениями первичных слуховых областей необходимо специальное исследование. Согласно результатам аудиометрического обследования с помощью длинных сигналов продолжительностью 1000-2000 мс, такие нарушения не приводят к какой-либо заметной потере слуха. Карп и др. путем измерения порогов восприятия длинных повторяющихся чистых тонов обнаружили ухудшение слуха в левом ухе, контралатеральном к пораженному правому полушарию, только у 10 из 19 пациентов с левосторонней гемиплегией, развившейся после инфаркта правого полушария мозга (Кагр et al., 1969). Эти данные не подтвердились в дальнейших исследованиях (Gershuni et al., 1971). Однако у пациентов с односторонними поражениями слуховой коры были выявлены серьезные трудности различения коротких сигналов продолжительностью 1-4 мс.
В наших исследованиях порог обнаружения белого шума измерялся с помощью слухового сигнала продолжительностью от 1 до 1000 мс. Использование короткого сигнала основывалось на наших предположениях об особой роли первич-
ных слуховых областей коры в переработке информации о сигналах малой продолжительности. В нашей первой публикации на эту тему (Бару и др., 1964) были представлены результаты исследования:
• 13 пациентов с инфарктами мозга;
• 8 пациентов с левыми верхнезадними височными поражениями;
• 5 пациентов с другими корковыми поражениями, в том числе двое пациентов с левыми теменными поражениями, двое пациентов с левыми нижними премоторными поражениями и один пациент с поражениями ствола мозга.
Было обнаружено, что пороги обнаружения белого шума продолжительностью 1000 и 100 мс левым и правым ухом у пациентов с левыми височными и левыми невисочными корковыми поражениями не различались. Снижение продолжительности сигнала до 1,5 и 4 мс привели к общему возрастанию порогов для левого уха до 26-28 Дб, как и у здоровых испытуемых. Пороги для левого уха увеличились в среднем до 36 Дб у пациентов с левыми верхнезадними височными поражениями, а у пациентов с невисочными корковыми поражениями остались на нормальном уровне 27-28 Дб (рис. 2.5.2). Средняя разность между порогами для правого и левого уха у пациентов височной группы составляла 9 Дб с разбросом от 6 до 26 Дб. В выборку также входил еще один пациент с инфарктом правой височно-затылочной области. У этого пациента пороги обнаружения белого шума продолжительностью 1000 мс для правого и левого уха не различались, а при уменьшении продолжительности сигнала до 1 мс пороги возросли, причем для левого уха на 9 Дб больше, чем для правого. Эти результаты указывают на большее возрастание порогов восприятия коротких слуховых сигналов для уха, контралатерального по отношению к пораженному полушарию. Аналогичные результаты обследования 20 пациентов с инфарктами левой верхнезадней височной области получены Вассерманом (Вассерман, 1968).
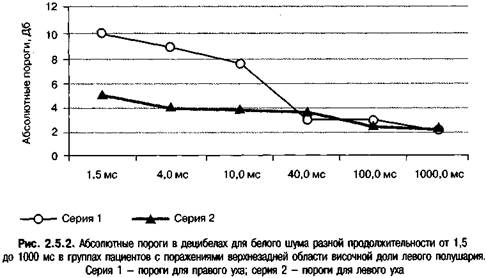
Карасева, Бару и др. наблюдали асимметричное возрастание порогов интенсивности коротких сигналов для уха, контралатерального к месту поражения, у 15 пациентов с поражениями правого полушария и 22 пациентов с поражениями левого полушария (Карасева, 1972; Бару и др., 1971). Большинство пациентов страдало опухолями височной доли. У девяти пациентов с поражениями слуховых структур ствола мозга, в том числе у одного пациента с проверенным поражением кохлеарных ядер и значительной потерей слуха, не было обнаружено асимметричного возрастания порогов для коротких звуков по сравнению с длинными звуками; пороги для длинных сигналов составляли 20-30 Дб. В шести случаях асимметричного возрастания порогов интенсивности для коротких звуков вскрытие показало поражения, затрагивающие кору и слуховые пути в районе извилины Гешля, а также ретроградную дегенерацию медиальных коленчатых тел. В двух проверенных случаях опухолей височных долей, где наблюдалась асимметрия для коротких звуков, эти области не были повреждены.
Эти данные указывают на роль А1, области первичной слуховой коры головного мозга человека, в переработке коротких слуховых сигналов, поступающих в ухо, контралатеральное к этой области. Аналогичные результаты были получены в исследованиях собак после односторонней резекции первичной слуховой области височной коры (Бару, 1966). Однако роль дегенеративного поражения медиальных коленчатых тел в описанных случаях не вполне ясна и требует дальнейших исследований.
Альберт и Бера описали пациента, который мог различать два щелчка только при увеличении межстимульного интервала (Albert, Bera, 1974). Лакнер и Тьюбер обнаружили увеличение времени слияния дихотически предъявленных щелчков у пациентов с поражениями левого полушария (Lackner, Teuber, 1973). Ауэрбах и др. получили те же результаты теста на слияние щелчков у пациента с полной словесной глухотой (Auerbach et al., 1982). Этот же пациент был не способен подсчитать количество щелчков, предъявляемых в быстром темпе.
По-видимому, выделение и распознание коротких сигналов является особенно важным в процессе распознания сложных свойств слуховых сигналов, которые часто включают в себя короткие составляющие частот и амплитуд. Требуются дальнейшие исследования этих взаимосвязей.
Дата добавления: 2015-09-18; просмотров: 1107;