СОСТАВ, СТРУКТУРНЫЕ ЭЛЕМЕНТЫ И СВОЙСТВА ДРЕВЕСИНЫ
Древесина состоит в основном из органических веществ, к которым полностью приложимы законы органической химии. Она является продуктом растительного происхождения и как биологический объект слагается из клеток. Стенки клеток древесины на 99% сложены из органических соединений, представленных у хвойных пород примерно на 70% углеводами, а у лиственных — на 80%. Углеводы, как известно, представляют группу природных веществ, образованных тремя элементами: углеродом, водородом и кислородом. К обширной группе этих веществ принадлежит сахар, а примером сравнительно простых углеводов может служить глюкоза (состав молекулы которой выражается формулой C6H12O6), иначе называемая моносахаридом. Молекулы простых Сахаров способны соединяться в растениях под влиянием ферментов в более крупные образования, например дисахарид: 2 C6H12O6 = C12H22O11 + H2O, а при большом количестве молекул моносахарида — в полисахариды, например с образованием крахмала (C6H10O5)n или целлюлозы с тем же выражением молекулы, но при более высоком значении n, чем у крахмала.
В углеводную часть древесины входит целлюлоза как ее основной компонент по объему в стволе дерева и нецеллюлозные полисахариды, именуемые гемицеллюлозами. Всю углеводную часть в целом называют холоцеллюлозой. Ее можно выделить из древесины в виде волокнистого материала обработкой окислителями (кислотами).
Около 30% древесины составляют вещества ароматической природы, известные под названием лигнина.
Углеводы и лигнин — это высокомолекулярные вещества, или полимеры. На них распространяются закономерности, характерные для этих соединений, причем они находятся в теснейшей взаимосвязи и образуют единую высокоорганизованную полимерную систему древесинного вещества.
К небольшой части древесины (2—4%) относятся экстрактивные вещества, которые не являются составляющими клеточной стенки. Они способны лишь пропитывать стенки клеток, а в основном содержатся в их полостях и в межклеточном пространстве. В отличие от углеводов и лигнина, экстрактивные вещества — низкомолекулярные соединения.
Следует несколько подробнее остановиться на характеристике главных структурных элементов древесины. Они, в совокупности с порами, капиллярами, контактными зонами и другими элементами микро- и макроструктуры, предопределяют основные свойства древесного материала.
Целлюлоза может быть выражена эмпирической формулой [C6H10O5]n или в другом виде [C6H7O2(OH)3]n, где n — степень полимеризации, которая у древесной целлюлозы достигает значений от 300 до 6000 и более. Она представляет собой линейный гетероцепной стереорегулярный[24] однородный полимер, имеющий большое число гидроксильных групп ОН, образующих водородную связь. Этот тип химической связи между водородным атомом гидроксила одной цепи и кислородным атомом гидроксила соседней цепи придает повышенную жесткость полимеру, так как «стягиваются», ориентируются цепные молекулы в целлюлозных волокнах.
Кроме водородных связей, для целлюлозы характерно также внутри- и межмолекулярное взаимодействие (т. е. силами Ван-дер-Ваальса), что, наоборот, уменьшает степень жесткости, и молекулы целлюлозы могут принимать различные конформации (расположения). Являясь основным веществом древесины, целлюлоза образует в структурном отношении слоистую клеточную оболочку (стенку), способную при механической обработке распадаться на тонкие целлюлозные волоконца-фибриллы, а при химической — на микрофибриллы. Фибрилла имеет кристаллическую структуру, так как для нее типичным является регулярное расположение молекул, характерное для молекулярной кристаллической решетки. Микрофибриллы также в основном сохраняют кристаллические (ориентированные) области. В некоторой части кристаллическая фаза перемежается с хаотическим (аморфным) расположением макромолекул, в ней отсутствует четко выраженная ориентация в микрофибриллах, а цепи значительно короче. Эту часть именуют гемицеллюлозой. Степень полимеризации макромолекул гемицеллюлозы составляет всего 100—200. Короткие цепи нередко попадаются среди кристаллических участков целлюлозного волокна, и тогда они достаточно прочно связываются с целлюлозой, образуя целлюлозаны, но оставаясь по существу гемицеллюлозой (ксиланы, маннаны).
Древесные целлюлозные волокна имеют спиральную структуру и содержат примерно 55—65% кристаллической и 25—35% аморф ной (гемицеллюлозной) части, причем у хвойных пород аморфной части меньше, у лиственных пород — больше (28—35%). Согласно другой теории, содержание кристаллической фазы в природной целлюлозе значительно больше, тогда как аморфной — не выше 5—10% и ее относят к дефектам упорядоченности. В этом случае полагают, что целлюлоза является однофазным кристаллическим веществом. Так или иначе, но главное, что необходимо учитывать, древесина — высококристаллическое вещество, имеющее форму продольных спиральных волокон, которые в деловой древесине выполняют роль своеобразного «армирующего» компонента, повышающего прочность материала на растяжение, чему способствует также их упорядоченное расположение.
Лигнин как другой структурообразующий компонент древесины — тоже природный полимер, представляет собой высокомолекулярную ароматическую часть, количество которой в древесине хвойных пород составляет 28—30%, а в древесине лиственных пород — 18—24%. В древесине он был открыт более 145 лет назад, но его весьма сложное строение до сих пор остается не полностью выясненным. Полагают, что это смесь нерегулярных разветвленных полимеров сетчатой структуры. Присутствие лигнина устанавливают по цветным реакциям. От целлюлозы отличается повышенным содержанием углерода — 60—65% по сравнению с 44% в целлюлозе, что обусловлено его ароматической природой. Из девяти атомов углерода, составляющих структурную единицу (фенилпропановую) лигнина, шесть принадлежат ароматическому кольцу. В химическом отношении лигнин — реакционно-способный полимер. По сравнению с целлюлозой лигнин обладает меньшей химической стойкостью, легче окисляется.
Лигнин — аморфное вещество; им обогащаются клеточные стенки с эффектом одревеснения. Он содержится в серединной пластинке клеточной стенки, но большая его часть находится во вторичной стенке (слое). Обнаруживается лигнин в этих двух слоях стенки по окрашиванию в красный цвет при воздействии на клеточные стенки химическим реагентом (солянокислородным флороглюцианом). Электронная микроскопия позволила, однако, не только по цветной реакции, но и на снимках увидеть, что лигнин заполняет также и межклеточные пространства. Своим присутствием он придает некоторую гидрофобность, но в целом она незначительна, и древесина относится к гидрофильным материалам, особенно древесина лиственных пород.
Оба основных компонента древесины — целлюлоза и лигнин — взаимодействуют между собой. Характер их взаимодействия еще полностью не раскрыт. Вначале предполагалось наличие чисто механической связи лигнина с углеводами в стенках (слоях) клетки. Такая теория называлась инкрустационной. Позднее было установлено, что невозможно ни извлекать углеводы из древесины без одновременного частичного удаления лигнина, ни полностью удалить лигнин из древесины без удаления некоторого количества углеводов. Очевидно, что это возможно только при их химическом взаимодействии, тем более что из древесины удалось выделить лигноуг- ' леводные комплексы. Большинство исследователей склоняется к предположению о существовании химической связи лигнина с геми-целлюлозой (ксиланом, маннаном и другими полисахаридами), хотя поддерживается мнение также о возможной химической связи лигнина с целлюлозой. В целом, однако, часть лигнина находится в древесине в свободном состоянии.
Экстрактивные вещества в отличие от главных компонентов извлекаются нейтральными растворителями — водой, обычными органическими растворителями. Хотя их немного, но они придают древесине цвет, запах, вкус, иногда токсичность, помогают дереву сопротивляться гниению, поражению грибами и пр. Среди экстрактивных веществ — смолы и смоляные кислоты, танниды (дубители), эфирные масла, красители, камеди, фитостерины, белки и пр. Содержание смол в лиственных породах до 1%, а в хвойных может быть до 20%. В каждой породе присутствуют только некоторые экстрактивные вещества, по-разному распределяясь внутри дерева, например фенольные вещества — в ядровой части, а сахара, жиры и др. — в заболонной древесине. Имеется небольшая доля и минеральных веществ (до 1%), поступающих из почвы через корневую систему и проводящие ткани.
Таким образом, древесина слагается из двух основных компонентов с присутствием небольших количеств других веществ, по-видимому, мало влияющих на формирование общей структуры.
Структура древесины в первом приближении представляет собой конструкционное сочетание целлюлозы с лигнином. Волокна целлюлозы обладают высокой прочностью на разрыв, но легко изгибаются (рис. 7.1). Лигнин объединяет их в единое целое с помощью водородных и ван-дер-ваальсовых сил связи и когезии, поэтому в совокупности древесина — по существу природный органический материал с конгломератным типом структуры, в котором имеются матричная пространственная сетка из лигнина и кристаллический волокнистый наполнитель в виде целлюлозы. Эта структура хорошо, например, видна на микроснимке, полученном американским ученым Э.Келли для среза осины при увеличении в 11000 раз (рис. 7.2). Темная полоса — лигнин, менее темная — стенка целлюлозной клетки и светлая область — полость клетки.
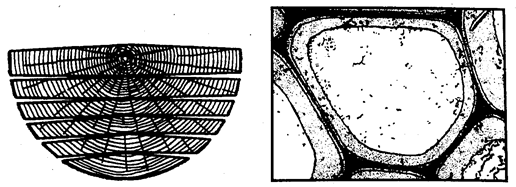
Рис. 7.1. Коробление древесины при высыхании (с появлением трещин усушки)
Рис. 7.2. Электронная микрофотография среза осины (11000 х по Э. Келли
Подобно искусственным, конгломератам, древесина содержит капилляры и поры различных размеров; в период жизни дерева они имеют большое значение для передвижения воды и питательных соков, их накопления и пр. Крупные капилляры являются полостями и порами стенок, капилляры и поры тонкие и мельчайшие находятся между фибриллами, микрофибриллами и внутри микрофибрилл. Если в крупных капиллярах и порах невелика удельная площадь внутренней поверхности, составляя, например, у ели 0,2 м2/г, то у мельчайших пор она очень большая, например у ели при диаметрах капилляров 10∙10-8—10∙10-7 см (от 10 до 100 А) удельная площадь их поверхности составляет до 300 м2/г. Такое различие капилляров отражается на характере контакта их с водой. Крупные капилляры могут заполняться водой, которая мало влияет на состояние древесины и ее качество (механические свойства) как строительного материала. Эта влага сравнительно легко приходит в капилляры и поры, особенно при контакте дерева с водой, заполняет полости и может составлять до 100—200% к массе абсолютно сухой древесины, но она сравнительно быстро и легко удаляется из них при сушке. Тонкие поры и капилляры заполняются не только при контакте с водой, но и в условиях влажного воздуха в связи с гигроскопичностью древесины и по законам капиллярных сосудов. Гигроскопическая влага сорбируется на стенках клеток, частично переходит в коллоидно-связанную с веществом дерева. Предельное насыщение древесины гигроскопической влагой составляет 25—35% (в среднем 30%) к массе абсолютно сухой древесины, называемое пределом насыщения. Насыщение гигроскопической влагой до этой предельной точки сопровождается набуханием древесины, изменением (ухудшением) ее физических и механических свойств. Увеличение влаги свыше 30%-ного ее содержания на механических свойствах древесины почти не отражается; не увеличивается и объем ее за счет набухания.
При сушке древесины трудно испаряется гигроскопическая (связанная) влага от клеточных стенок насыщения- волокон, причем объем древесины уменьшается в связи с усадкой и уплотнением клеточных оболочек. Особенно трудно удаляются последние 4—6% гигроскопической влаги, так как она ориентированно закреплена (адсорбирована) в монослое молекулами целлюлозы. Возникают водородные связи между гидроксилами целлюлозы и водой, тогда как другая большая часть гигроскопической влаги (20—25%) находится под влиянием капиллярной конденсации. С приближением при сушке к температуре 105°С масса древесины сохраняет постоянное значение, что и принимают за абсолютно сухое состояние древесины. Абсолютно сухая древесина состоит из Двух компонентов — древесинного вещества и воздуха. Фактически около 1% гигроскопической влаги в абсолютно сухой древесине сохраняется, но за счет относительно прочных водородных связей она не снижает ее качественных показателей. Дальнейшее повышение температуры вызывает более или менее глубокое разложение целлюлозы — деструкцию, особенно в присутствии воздуха и влаги. При умеренном нагревании (120—180°С) изменяется цвет целлюлозы, снижается ее прочность, а при более высокой температуре (230—240°С) протекают химические реакции с изменением элементарного состава целлюлозы. При температуре, близкой к 300°С,структура переходит из кристаллической формы в аморфную, а при дальнейшем повышении температуры образуются целлюлозный уголь и жидкие продукты распада (уксусная кислота, ацетон, формальдегид, муравьиная кислота и др.). Лигнин более устойчив к термолизу, чем целлюлоза, за счет его ароматического строения. Однако и он к моменту обугливания целлюлозы подвержен экзотермическому распаду с потерей своих первоначальных свойств. Следовательно, при нагревании выше 105—110°С непрерывно протекают процессы деструкции, а древесина сравнительно быстро теряет свои высокие физико-механические свойства, которые она имела в абсолютно сухом состоянии.
Кинетику изменения показателей свойств древесины по мере ее высыхания можно изобразить графически в системе прямоугольных координат. Для этого на параллельно расположенных осях ординат необходимо отложить в соответствующих масштабах различные показатели свойств (на рисунке условно принята одна ось ординаты для всех свойств), а на оси абсцисс — влажность, еще лучше структурный параметр в виде, например, отношения объема V свободной влаги, равного MВ/ρВ или гигроскопической MГ.В/ρВ отнесенных к суммарной площади стенок пор и капилляров, постоянной для данной породы дерева. Очевидно, что отношение V/Aсум = δ, где δ — усредненная толщина пленки свободной или гигроскопической влаги; ρВ — плотность воды.
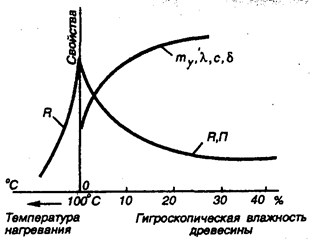
Рис. 7.3. Проявление закона створа в древесине при отсутствии влаги:R — прочность, mν — средняя плотность, λ —теплопроводность, с — электропроводность, δ — толщина пленки воды, П — пористость
По мере испарения свободной влаги в свежесрубленной древесине, когда ее влажность составляет обычно от 40 до 100% и выше, показатели свойств остаются постоянными до предела насыщения волокон. При постепенном высушивании в пределах содержания гигроскопической влаги с утоньшением полимолекулярных слоев (δ) воды на развитой поверхности тончайших капилляров и пор показатели свойств непрерывно улучшаются. К моменту высыхания, когда влажность становится равной 1% и меньше, а гигроскопическая вода достаточно прочно химически связана с целлюлозой в монослоях, причем отношение V/Асум принимает минимальное значение, близкое к нулю, тогда показатели свойств становятся экстремальными: прочность, выражающая любые ее значения (при сжатии, растяжении и т. п.), — наибольшей; упругость — также наибольшей, тогда как пластичность и общая деформация — наименьшими, теплопроводность и электропроводность — наименьшими (в сухом состоянии древесина является хорошим диэлектриком); пористость (П), коэффициент диффузии при постоянной температуре — наибольшими, плотность — наименьшей и т. д. (рис.7.3). Числовые значения этих экстремумов получают экспериментальным путем при испытании образцов или расчетным, применяя некоторые известные зависимости. В них значение влажности для получения экстремальной величины принимается равным нулю (W = 0).
На левой ветви получаемой экстремальной кривой показаны значения свойств по мере развития деструкционных процессов при дальнейшем повышении температуры древесины (свыше 105°С).
Из графика следует, что при определенном наборе структурных параметров в абсолютно сухой древесине закономерно возникает комплекс экстремальных значений тех физических и механических свойств, которые непосредственно связаны со структурой. Такой комплекс экстремумов согласно закону створа возможен только при оптимальной структуре.
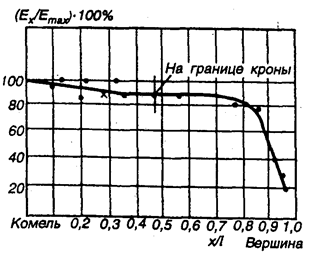
Рис. 7.4. Результаты экспериментального определения модуля упругости Е при изгибе древесины сосны на растущем дереве (по Юлинену): х — расстояние от комля; l — высота ствола
Следовательно, под влиянием внешних и внутренних детерминантов в период роста дерева происходит постепенная оптимизация структуры древесины эволюционным путем. К внешним, воздействовавшим на растущее дерево, следует отнести: силовые — от ветровой нагрузки и массы кроны, утяжеленной атмосферными осадками; тепловые — от переменной температуры окружающей среды; воздействие влаги, солнечного света и др. К внутренним детерминантам относятся: перемещение влаги с растворенными минеральными веществами из почвы; перемещение и создание запаса питательных веществ; процесс фотосинтеза в кроне и др. Реальные условия развития дерева не только благоприятствовали его росту, но и вызывали необходимость в приспособляемости к механическим напряжениям, особенно древесных волокон. Так, сопротивляемости ветровым нагрузкам благоприятствовало: развитие слоистого строения древесины с правильным чередованием прослоек более мягкой весенней и более жесткой летней древесины; повышение модуля упругости древесины ствола от вершины к комлю (основанию) (рис. 7.4), сбег толщины ствола от комлевой части его к вершине при высокой прочности на растяжение корней. Прочные наружные слои ствола при меньшей прочности сердцевины обеспечивают дереву надежную сопротивляемость большим вертикальным нагрузкам от массы кроны. В результате нормального роста древесина хвойных и лиственных пород набирает комплекс анатомических элементов, общим для которых является упорядоченность волокнистой структуры и клеточного строения древесины, утончаются стенки-прослойки матричной части из лигнина, наступает в процессе значительной кристаллизации целлюлозы и, отчасти, гемицеллюлозы общая упорядоченность расположения молекул с анизотропией свойств по главным структурным направлениям, формируется оптимальная структура с соответствующей закономерностью створа с комплексом экстремальных значении свойств. В этом проявляется структурная детерминация развития растений.
7.3. АНАТОМИЧЕСКОЕ СТРОЕНИЕ ДРЕВЕСИНЫ
Для полной характеристики древесины и древесных строительных материалов следует раскрыть еще одну сторону структуры и свойств этого биологического объекта растительного происхождения: взаимосвязь анатомического строения с физико-химическими и физическими свойствами древесины, тем более что здесь прослеживаются различия между хвойными и лиственными породами, как на уровне микро-, так и макроструктуры. Принято изучать три основных разреза ствола (рис. 7.5).
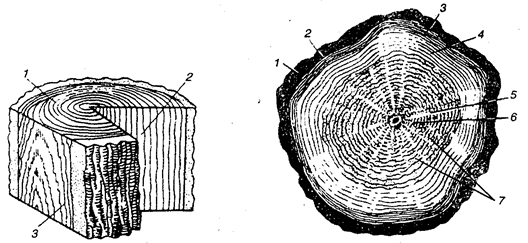
Рис. 7.5. Основные разрезы ствола: 1 — торцовый; 2 — радиальный; 3— тангенциальный
Рис. 7.6. Торцовый разрез ствола: 1 — кора; 2— луб; 3— камбий; 4— заболонь; 5 — ядро; 6 — сердцевина; 7 — сердцевинные лучи
В торцовом разрезе ствола дерева выделяют (рис. 7.6): сердцевину (сердцевинная трубка) — примерно в центре ствола размещается вдоль его оси, которая является рыхлой первичной малопрочной тканью; собственно древесину, слагаемую из концентрически расположенных годичных слоев с включением в каждый из них весенней и более плотной летней древесины; камбий — тончайший слой из полностью живых клеток, способных к росту и делению на большую часть, откладываемую в сторону древесины, и меньшую — в сторону от центра, где расположен следующий слой в виде луба; луб является внутренним слоем коры (флоэмы) и граничит с внешним слоем ее, называемой коркой (темного цвета). На долю ствола приходится 70—90% всего объема дерева; остальную часть составляют крона и корневая система. Кора составляет от 6 до 25% объема ствола, остальная часть его служит древесиной, в которой наблюдается светлое периферическое кольцо — заболонь и более темная центральная часть — ядро. Камбиальный слой находится между заболонью и лубом. Все эти части составляют макроструктуру древесины. У некоторых древесных пород (дуб, бук, клен и др.) на торцовом сечении можно видеть узкие радиально расположенные полоски, которые называются сердцевинными лучами 7.
В породах может отсутствовать ядровая часть, и тогда породы именуются заболонными (береза, липа, клен, граб и др.). В других породах заболонь имеет цвет центральной части ствола, причем последняя остается более сухой; такие породы называются спелодревесными (ель, пихта, осина, бук и др.).
Более подробное строение древесины хвойных и лиственных пород изучается с помощью микроскопа и специальных срезов слоев древесины.
Выше отмечалось, что древесина состоит в основном из клеток. Их можно разделить на два типа: прозенхимные и паренхимные. Прозенхимные имеют длину во много раз большую, чем их ширина; они придают древесине волокнистое строение, оставаясь, как правило, уже отмершими. Среди этих клеток различают трахеиды, клетки либриформа и сосуды. Паренхимные. клетки — короткие, имеют примерно одинаковую длину и ширину, являются живыми. Если в живой клетке имеется оболочка (клеточная стенка), внутри которой расположены протоплазма (протопласт) и ядро, то в отмершей в процессе роста дерева протопласт частично расходуется на утолщение стенки, частично высыхает вместе с ядром, оставляя полость, которая заполняется водой или воздухом, а иногда экстрактивными веществами. Естественно, что в срубленном дереве живые клетки быстро умирают, и оно состоит из омертвевших клеток, т. е. из клеточных стенок (оболочек).
Клетки в древесине имеют различное функциональное значение. Одни из них выполняют функции проводящих клеток, другие — опорных или механических, третьи — запасающих, четвертые — образовательных (расположены в камбиальном слое), пятые — ассимиляционных (находятся в листве и хвое и благоприятствуют образованию питательных веществ), шестые — покровных (в корке дерева).. Клетки, имеющие одинаковое строение и выполняющие одну и ту же функцию, образуют ткани. Особо выделяют ткани проводящие, механические и запасающие, что соответствует первым трем функциям их клеток.
Древесина хвойных пород имеет относительно простое строение (рис. 7.7). Она состоит из клеток почти одного типа — трахеид (рис. 7.8). К ним относятся мертвые веретенообразные клетки длиной от 1,5 до 5 мм со стенками разной толщины и с полостями различных размеров. Трахеиды весенней древесины имеют широкие полости и тонкие стенки, а осенней — более узкие полости и толстые стенки. У весенних трахеид, выполняющих функции водопро-водящих клеток, отношение диаметра к длине составляет 1:100, у осенних — 1:400. В стенках трахеид имеются поры, через которые клетки общаются между собой и при помощи которых содержимое живых клеток соединяется в одно целое.
Паренхимные клетки составляют живую ткань хвойной древесины; находятся, главным образом, в сердцевинных лучах и, следовательно, выполняют функции запасающих питание (крахмал и жиры) для потребления весной, а также в небольшом количестве в лубе, поблизости от камбия (вертикальная паренхима). Кроме того, паренхимные клетки, образуя межклеточные каналы, как бы выстилают поверхность смоляных ходов, столь характерных для хвойных. пород. Такие ходы, располагаясь вертикально и горизонтально, образуют единую смолоносную систему, что в конечном итоге благоприятствует повышению стойкости древесины хвойных пород. В целом, однако, объемная доля паренхимной ткани в хвойной древесине составляет 3—5%, так как часть запасов питания хранится в хвое, где не имеется паренхимных клеток.
Лиственные породы имеют более сложное анатомическое строение (рис. 7.9). Проводящими (водопроводящими) клетками (элементами) служат сосуды длинные трубки шириной 0,02—0,5 мм (рис. 7.10). В весенней древесине они более широкие и их можно иногда заметить невооруженным глазом. Стенки сосудов характерны утолщением кольчатой, спиральной или сетчатой формы, что придает стенкам повышенную прочность. Механические клетки, и соответственно механическая ткань, называемые у лиственных пород либриформом, заметно отличаются своей веретенообразной формой, толстыми стенками с щелевидными порами, узкими полостями клеток, имеют сравнительно небольшие длину (0,3—0,8 мм) и диаметр (0,1—0,2 мм). Все клетки либриформа являются мертвыми, и лишь паренхимные клетки как и в хвойных породах, образуют сердцевинные лучи, как запасающие и проводящие питательные вещества в радиальном направлении при объемной доли их около 10%, т. е. в 2—3 раза больше, чем в хвойных породах.
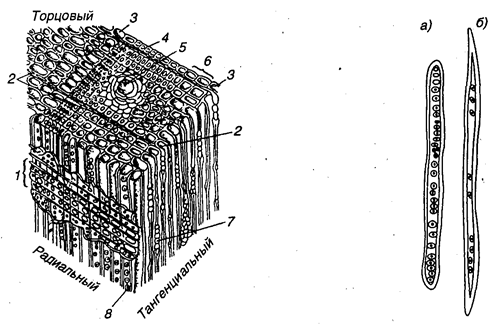
Рис. 7.7. Микроструктура древесины сосны: 1 — сердцевинный луч в радиальном разрезе; 2 — многорядный луч; 3 — годичный слой; 4 -поздние трахеиды; 5 — вертикальный смоляной ход; 6 —ранние слои древесины; 7 — сердцевинный луч в тангенциальном разрезе; 8 — окаймленная пор
Рис. 7.8. Трахеиды ранней (а) и поздней (б) древесины сосны
Выше были отмечены шесть функций клеток, но при кратком описании анатомического строения древесины были описаны лишь три из них, хотя три других являются не менее значимыми. Образовательная функция — живые клетки камбия образованы тонкими оболочками и наполнены протоплазмой и ядром. В процессе роста древесины клетка камбия делится на две неравные части с возникновением одной новой активной камбиальной клетки; вторая становится либо клеткой древесины, либо — луба. В обоих случаях после отделения клетки от камбиального слоя в ней начинается лигнификация с отложением лигнина, повышением ее гидрофобности и одревеснения, затем она отмирает. Процесс роста древесины неравномерный, что легко устанавливается по плотности и ширине годичных слоев (годичных колец): поздняя древесина (осенняя) — плотнее и темнее весенней, а ширина слоев зависит от породы древесины. При необходимости по этим признакам можно определить процент содержания поздней древесины; чем он выше, тем прочнее древесина как строительный материал.
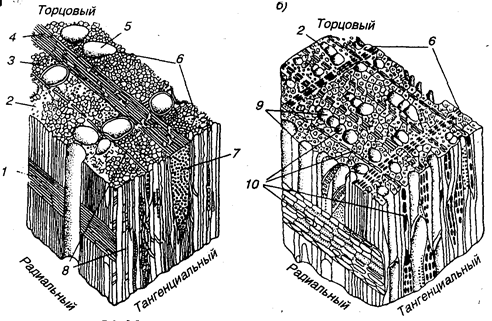
Рис. 7.9. Микроструктура древесины лиственных пород:
а — дуба; б — березы; 1 — сердцевинный луч; 2 — либриформ; 3 — мелкие сосуды; 4 — широкий сердцевинный луч в торцовом (поперечном) разрезе; 5 — крупный сосуд; 6 — годичный слой; 7 — широкий сердцевинный луч в тангенциальном разрезе; 8 — узкий сердцевинный луч; 9 — сосуды; 10 — сердцевинные лучи в различных разрезах

Рис. 7.10. Сосуды лиственных пород:1 — липы; 2 — бука; 3 — дуба
Ассимиляционная функция клеток заключается в усвоении клетками зеленых листьев и хвои внешних веществ (воды, углекислоты) с образованием углеводов в результате фотосинтеза, т. е. при помощи лучистой (солнечной) световой энергии, поглощаемой хлорофиллом. Последний находится в виде хлорофилловых зерен в клетках листа, являясь в нем красящим веществом, или пигментом, придающим листьям зеленую окраску. Сущность процесса ассимиляции раскрыта великим русским ученым К.А. Тимирязевым (1843—1920). Ее схематически можно представить в следующем виде (рис. 7.11): листья поглощают из атмосферы углекислый газ СО2; хлорофилловые зерна в листьях на солнечном свету поглощают продиффундировавший углекислый газ СO2 и выделяют кислород в воздух; оставшийся в листьях углерод вступает во взаимосвязь с молекулами воды, проникшей к кроне дерева из почвы по сосудам заболонного слоя древесины; в результате такого взаимодействия образуется промежуточное вещество — формальдегид СH2O; шесть молекул газообразного формальдегида (с резким неприятным запахом и вкусом) в результате фотосинтеза и полимеризации (т. е. соединения друг с другом) образуют новую молекулу вещества, именуемого глюкозой: СH2O = С6Н12О6; глюкоза из листьев по сосудам переходит в различные части древесины и отлагается там в запас; под влиянием молекул воды и ферментов n молекул глюкозы переходят в одну молекулу крахмала, а именно: nС6Н12О6 = nH2О + (C6H10O5)n Величина индекса n — степени полимеризации крахмала — высокая, но не до конца пока выясненная по своему числовому значению. Однако она может оказаться еще больше с возникновением макромолекулы целлюлозы (клетчатки) со степенью полимеризации до 6000 и выше. Из таких макромолекул целлюлозы в процессе роста дерева слагаются стенки клеток древесины.
Немаловажную роль в росте и сохранности дерева имеют функциональные клетки — покровные. Они возникают при делении клеток камбия с отложением их в сторону луба, а затем полностью отмирают и переходят, как структурообразующий элемент, во внешний слой коры дерева, т. е. в корку. Корка предохраняет дерево — древесину, камбиальный слой и луб (проводящий питательные вещества от кроны в ствол и корневую систему) — от механических повреждений, вредного воздействия внешней среды, низких температур или резких температурных колебаний.
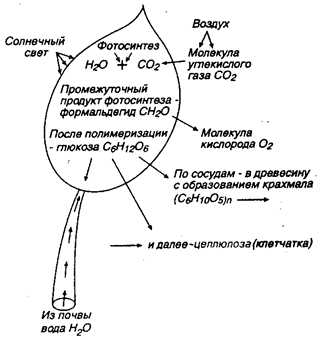
Рис. 7.11. Схема ассимиляции с фотосинтезом в зеленых листьях дерева (по К.А. Тимирязеву)
Дата добавления: 2015-09-18; просмотров: 1991;