Обмен жирных кислот.
Жирные кислоты могут вступать в реакции только после активации. Активация жирных кислот происходит в цитоплазме. Для активной жирной кислоты, как и для глицерина, возможны два пути метаболических превращений: 1. Катаболизм до Ацетил-КоА. Этот процесс называют b-ОКИСЛЕНИЕМ ЖИРНЫХ КИСЛОТ. 2. Синтез жира или других липидов.
B - ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ. РЕГУЛЯЦИЯ b-ОКИСЛЕНИЯ
Жирные кислоты, проникающие из крови в клетку, сначала подвергаютсяреакции активации под действием фермента ацил-КоА-синтетазы:
RCOOH+HSKoA+ATP ® R-CO-SKoA+AMP+PPi
B-окисление жирных кислот — это специфический путь распада жирных кислот, заканчивающие образованием ацетил-КоА. b-окисление жирных кислот имеет такое название потому, что реакции окисления в радикале жирных кислот происходит по b--углеродному атому.b-окисление жирных кислот и последующее за ним окисление ацетил-КоА в ЦТК служатисточником энергии для синтеза АТР.
Процесс b-окисления происходит в матриксе митохондрий и только ваэробных условиях, так как связан с ЦПЭ. |
Внутренняя мембрана митохондрий непроницаема для ацил-КоА, поэтому существует система переноса жирных кислот через мембрану в комплексе с молекулой карнитина (рис.9)
Во внешней мембране митохондрий находится фермент карнитинацилтрансфераза I, который катализирует перенос ацила с КоА на небольшую молекулу карнитина. Затем ацилкарнитин с помощью транслоказы переносится через внутреннюю мембрану митохондрий, где фермент карнитинацил-трансфераза II переносит ацил на внутримитохондрильный HSKoA.
После того как ацил-КоА попадает в матрикс митохондрий, начинается процесс b-окисления, представляющий собой 4 последовательные реакции, которые заканчиваются укорочением жирной кислоты на 2 углеродных атома, так как отщепляется ацетильный остаток (рис.10).
Эти 4 последовательные реакции повторяются до тех пор, пока вся жирная кислота, имеющая четное число атомов углерода, не превратится в определенное количество молекул ацетил-КоА. Эти 4 реакции b-окисления (дегидрирование, гидратация, дегидрирование, отщепление ацетил-КоА) обычно называют циклом b--окисления, так как имеется в виду, что одни и те к реакции повторяются с радикалом жирной кислоты до тех пор, пока вся кислота не превратится в ацетильные остатки.
Количество молекул АТР, которые образуются при окислении жирной кислоты, можно точно рассчитать. Для этого необходимо знать, что в каждом цикле:
а) образуется ацетил-КоА, который в ЦТК окисляется до СО2 и воды.
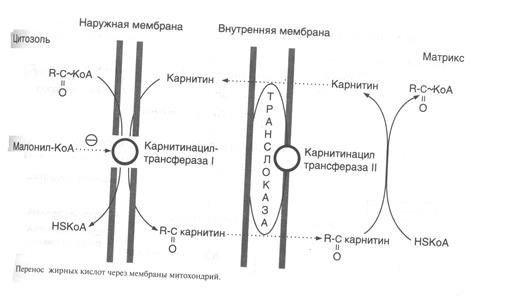
Рисунок 9
Число молекул ацетил-КоА, образующихся в результате окисления жирной кислоты с числом п атомовС, можно рассчитать по формуле: п / 2,(п / 2) х 12 = количество молекул АТР,
б) при b--окислении происходят 2 реакции дегидрирования, в которых восстанавливаются 1 молекула убихинона и 1 молекула NAD+, поэтому каждый цикл дает 5 молекул АТР с участием ЦПЭ;
в) число циклов можно рассчитать по формуле: (п / 2) – 1, так как в последний цикл b--окисления всегда вступает бутирил-КоА и при его окислении образуется 2 ацетил-КоА, а не один, как во всех предыдущих циклах;
г) суммарный выход АТР при окислении жирной кислоты с числом п атомов С можно рассчитать по формуле:
[ {(п / 2) х 12} + {(п / 2) – 1)} х 5] – 1* = число молей АТФ/ моль жирной кислоты.
*1 молекула АТР используется на активацию жирной кислоты.
Например, при окислении пальмитиновой кислоты (С16) происходит 7 циклов b-окисления, в результате которых образуется 8 моль ацетил-СоА, 7 моль FADH2 и 7 моль NADH+Н+. Следовательно, выход АТР составляет 35 АТР в результате b-окисления и 96 АТР в результате цитратного цикла, что соответствует в сумме 131 моль АТР.
Регуляция b-окисления. Скорость b-окисления, так же как и других метаболических путей, зависит от доступности субстрата ацил-КоА, поэтому b-окисление жирных кислот активируется в постабсорбтивный период или при длительной физической работе, когда в результате распада жиров в жировой ткани в крови увеличивается концентрация жирных кислот. В этих условиях мышцы, миокард и печень активно используют жирные кислоты как источник энергии. Мозг не использует жирные кислоты как источник энергии, так как они не проникают через гематоэнцефалический барьер, являясь гидрфобными молекулами.
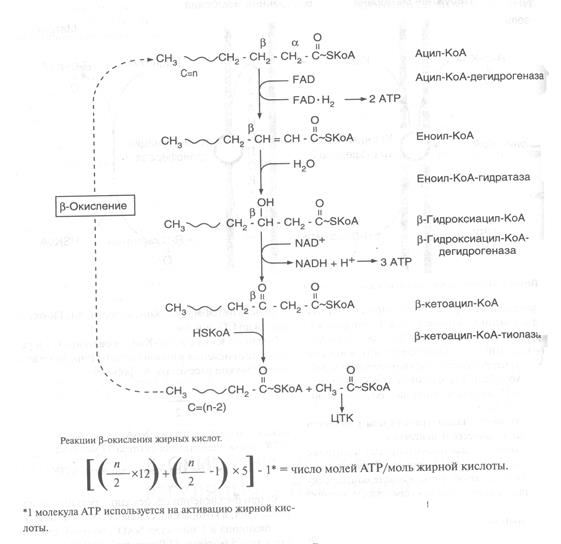
Рис. 10
Регуляторный фермент b-окисления - карнитинацилтрансфераза I. Аллостеричный ингибитор этого фермента — малонил-КоА образуется только при биосинтезе жирных кислот, следовательно, в постабсорбтивный период, когда поступление ацетильных остатков из митохондрий в цитозоль прекращается, синтез малонил-Ko тоже прекращается иb-окисление в отсутствие ингибитора активируется.
Как важнейший путь, поставляющий АТФ, b-окисление активируется при увеличении в клетке потребности в энергии. Это возможно благодаря непосредственной связи реакций b-окисления через коферментыNAD и FAD с цепью переноса электронов. Чем интенсивнее идет распад АТФ, тем быстрее окисляются жирные кислоты, обеспечивая синтез новых молекул АТФ.
Дата добавления: 2015-09-18; просмотров: 1394;