Подовження кінців еукаріотичної хромосоми
Ще одна характерна відмінність еукаріотичної хромосоми полягає в тому, що вона, на відміну від прокаріотичної, є лінійною – має два кінці. Унаслідок цієї простої обставини на 3′-кінцях матричних ланцюгів ДНК залишаються одноланцюгові хвости (рис. 21): два РНК-праймери на 5′-кінцях синтезованих ланцюгів видаляються, а прогалина не може бути заповненою, оскільки немає 3′-кінця, який міг би бути використаним як праймер. Одноланцюгові хвости піддаються швидкій нуклеазній деградації і після кожної реплікації хромосома має вкоротитися.
Кінцеві ділянки ДНК еукаріотичної хромосоми – теломери – складаються з невеликих елементів послідовності, що тандемно повторюються – теломерних повторів. Подовження теломер після реплікації здійснюється за допомогою спеціального ферменту – теломерази, яка є РНК-залежною-ДНК-полімеразою. РНК-матриця входить до складу самого ферменту і містить ділянку, комплементарну теломерному повтору (рис. 22). Використовуючи цю ділянку як матрицю і 3′-кінець як праймер, теломераза покроково добудовує до 3′-кінця кілька копій теломеразного повтору. Далі подовжений одноланцюговий хвіст використовується як матриця для синтезу іншого ланцюга за звичайним реплікативним механізмом. Видалення РНК-праймера після цього не є проблемою, оскільки хромосома вже є подовженою.
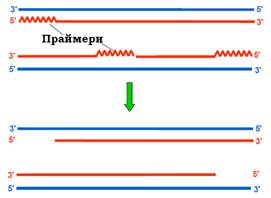
Рис. 21. Дві дочірні лінійні молекули ДНК після реплікації.
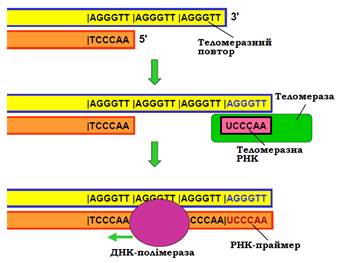
Рис. 22. Подовження кінців хромосоми за допомогою теломерази.
Теломераза є активною в клітинах, що розвиваються, і злоякісно-трансформованих клітинах і неактивною – у диференційованих соматичних клітинах вищих еукаріотів. Відповідно, певне критичне скорочення теломерів, яке відбувається в таких клітинах після кількох десятків клітинних поділів, є одним із механізмів активації програми їхньої загибелі.
Дата добавления: 2015-09-11; просмотров: 957;