Гомологічна рекомбінація
Необхідною умовою для здійснення гомологічної рекомбінації є загальна гомологія між двома молекулами ДНК по всій довжині. Загальну модель початкового етапу гомологічної рекомбінації зображено на рис. 1.
Ініціюючою подією є дволанцюговий розріз в одній з гомологічних молекул. Цей розріз розширюється шляхом 5′-екзонуклеазної деградації ДНК – у результаті в місці розрізу залишаються два одноланцюгові 3′-хвости. Один із них здійснює інвазію – утворює подвійну спіраль з антипаралельним ланцюгом інтактної гомологічної молекули ДНК. Інший ланцюг цієї останньої, відповідно, виштовхується з дуплекса у вигляді одноланцюгової D-петлі. D-петля (від displacement), разом із 3′-хвостом, здатна переміщуватись у пошуку гомології – максимальної комплементарності в межах подвійної спіралі, що утворилася між ланцюгами двох гомологічних молекул ДНК. На наступному кроці відбувається репараційний синтез ДНК: два 3′-кінці розірваної молекули ДНК подовжуються ДНК-полімеразами з використанням у ролі матриць двох ланцюгів інтактної молекули. До цього моменту схема на рис. 1 є одночасно схемою точної репарації дволанцюгового розриву в одній із двох сестринських молекул ДНК під час реплікації.
Під час рекомбінації відновлення цілісності ДНК є тільки завершенням початкового етапу. У результаті інвазії та репараційного синтезу дві молекули ДНК об’єднуються в чотириланцюгову структуру з двома перехрестями ланцюгів – структурами Холідея (Robin Holliday). Кожна структура Холідея може переміщуватись (так звана міграція гілки), результатом чого є подовження гетеродуплекса – подвійної спіралі між двома майже комплементарними ланцюгами двох гомологічних молекул ДНК.
Вважається, що за схемою на рис. 1 відбуваються початкові стадії гомологічної рекомбінації в більшості про- та еукаріотів. Але механізми цих процесів найкраще вивчені для бактерій. В E. coli два кінці розірваної молекули ДНК упізнаються комплексом трьох білків – recBCD (рис. 2), який має геліказну та екзонуклеазні активності. Комплекс починає рухатися, руйнуючи подвійну спіраль і одночасно деградуючи ДНК за рахунок своїх 5′- та 3′-екзонуклеазних активностей. У так званому χ-сайті (сайт підвищеної частоти рекомбінації в E. coli – послідовність довжиною вісім пар основ, що зустрічається із середньою періодичністю у 5 тис. пар основ) 3′-екзонуклеазна активність гальмується, результатом чого є утворення одноланцюгового 3′-кінцевого хвоста. Цей хвіст відразу вкривається білком recА. Останній має два сайти зв’язування з ДНК, за рахунок першого з них і відбувається (у присутності АТР як кофактора) кооперативна взаємодія з одноланцюговою ДНК: молекули білка оточують полінуклеотидний ланцюг, формуючи на його поверхні праву спіраль. Саме цей комплекс і здійснює інвазію в подвійну спіраль інтактної гомологічної молекули ДНК: другий сайт взаємодії recА з ДНК використовується на цій стадії для утворення комплексу з двома ланцюгами гомологічних молекул ДНК. За умови комплементарності двох ланцюгів здійснюється гідроліз АТР, після чого recА втрачає спорідненість до ДНК, залишаючи сформований гетеродуплекс.
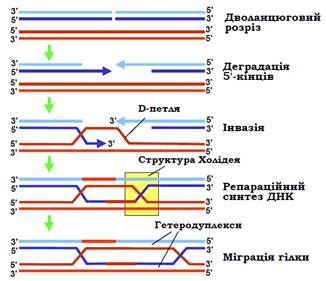
Рис. 1. Початкові стадії гомологічної рекомбінації.
Повертаючись до останньої панелі на рис. 1, розглянемо окрему структуру Холідея. Як показано на рис. 3, її можна піддати ізомеризації, повернувши два дволанцюгові кінці на 180°. Саме у формі без перехрестя ланцюгів (праворуч на рис. 3) структура Холідея фіксується завдяки її взаємодії з гомотетрамерним білком ruvA: білок зв’язується з центром хреста, утримуючи чотири одноланцюгові ділянки у приблизно планарній квадратній конфігурації (рис. 4, а). З ruvA та двома дуплексами, що виходять з хреста у протилежних напрямках, взаємодіє білок ruvВ (шість субодиниць, які оточують подвійну спіраль кільцем). RuvВ працює як АТР-залежна геліказа (або, швидше, як комплекс ремоделювання хроматину: два гексамери ruvВ генерують обертальні рухи подвійної спіралі у протилежних напрямках, що приводить до протягування ланцюгів через комплекс ruvА/ruvВ (рис. 4, б). Саме активність ruvВ і забезпечує переміщення структури Холідея – міграцію гілки з одночасним подовженням гетеродуплекса.
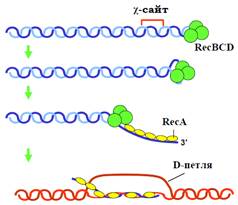
Рис. 2. Механізм початкових стадій гомологічної рекомбінації в E. Coli.
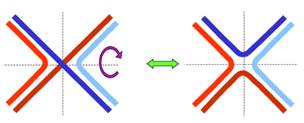
Рис. 3. Ізомеризація структури Холідея.
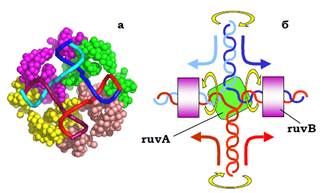
Рис. 4. (а): Комплекс ruvA зі структурою Холідея (1BDX, показано лише С α-атоми білка та Р-атоми ДНК).(б): Схема переміщення структури Холідея за рахунок активності ruvB: стрілки вказують напрямок обертання дуплексів та напрямок руху ланцюгів через комплекс ruvA/ruvВ
(колір стрілок збігається з таким ланцюгів).
Зі схеми на рис. 5 видно, як можна ототожнити протягування чотирьох полінуклеотидних ланцюгів із міграцією гілки. На схемі у два етапи проведено формальну ізомеризацію планарної конфігурації структури Холідея в конфігурацію з перехрестям: на першому етапі два ланцюги у складі хреста випрямляються, на другому – нижня частина отриманої конфігурації обертається на 180°. Очевидно, що зображене на рис. 4, 5 протягування ланцюгів через комплекс ruvA/ruvВ є еквівалентним зсуву перехрестя у правий бік.
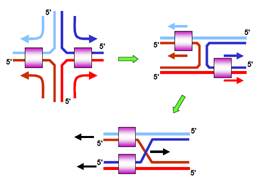
Рис. 5. Спрощена схема комплексу ruvВ зі структурою Холідея з рис. 4 та її формальні перетворення у дві топологічно еквівалентні конфігурації. Остання конфігурація збігається з правою структурою Холідея
з рис. 1. Кольорові стрілки вказують напрямок руху ланцюгів через ruvВ,
чорні – напрямок руху дуплексів і перехрестя.
Останньою подією рекомбінації є розділення (resolution) структури Холідея резольвазою – білком ruvС. Резольваза є ендонуклеазою, дві молекули якої взаємодіють з комплексом ruvА/ruvВ і двома ланцюгами хреста, розташованими один навпроти одного: є два рівноймовірні варіанти такого зв’язування. Отже, резольваза робить дволанцюговий розріз через хрест двома можливими шляхами (рис. 6). Після наступного зшивання розривів лігазою залишаються дві дволанцюгові молекули ДНК.
На рис. 6 показано ізомеризовану чотириланцюгову структуру з рис. 1 – дві структури Холідея з перехрестями перетворені на планарні (ділянки, синтезовані шляхом репарації, не позначено) – і результат її розділення. Із чотирьох можливих комбінацій розділення двох структур Холідея показано дві. Одна з цих комбінацій викликає рекомбінацію (аналогічно, рекомбінантною є і пара розрізів 2 + 4): дві гомологічні молекули ДНК обмінялися ділянками й умовна «абетка», якою позначено фрагменти ДНК, змінює регістр – великі літери замінюються на маленькі й навпаки. Пари розрізів 1 + 4 і 2 + 3 не приводять до рекомбінації. Таким чином, рекомбінація при розділенні структур Холідея відбувається з імовірністю 50%.
Незалежно від того, чи відбулася рекомбінація, усі продукти містять гетеродуплекси (середня частина всіх кінцевих молекул на рис. 6). Оскільки гетеродуплекси складаються з майже комплементарних ланцюгів, вони містять місметчі. Відповідно, кінцевою операцією, яка завершує процес гомологічної рекомбінації, є репарація цих місметчів системою mutHLSU. На відміну від того, як ця система спрацьовує після реплікації, після рекомбінації ланцюг, де відбувається заміна нуклеотидів, обирається випадково. Наприклад, центральний фрагмент першої молекули нарис. 6 перетворюється або на В/В′, або на b/b′. Якщо на цій ділянці розташований ген, шляхом репарації обирається один із його алелів – В або b. Отже, побічним ефектом рекомбінації є відоме в генетиці явище конверсії – гена. Білки системи mutHLSU якимось чином блокують рекомбінацію на ранніх етапах, коли місметчів виявляється надто багато (так звана гомеологічна рекомбінація): міграція гілки зупиняється і чужорідна ДНК виштовхується з гетеродуплекса.
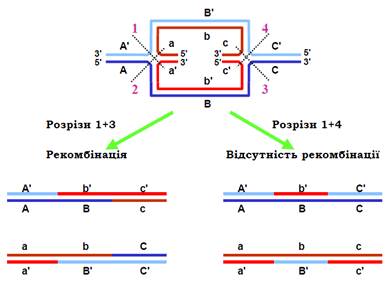
Рис. 6. Схема розділення двох структур Холідея. Угорі: конфігурація чотирьох ланцюгів еквівалентна конфігурації на нижній панелі з рис. 1. Літерами позначено ділянки ланцюгів (великі й маленькі літери відповідають гомологічним ділянкам двох молекул, літери зі штрихом і без – комплементарним ділянкам вихідних дуплексів). Цифрами 1 – 4 позначеноможливі розрізи резольвазою. Унизу: дві пари дволанцюгових молекул після розділення структур Холідея, отримані в результаті відповідних розрізів.
Еукаріотичні клітини містять білки, гомологічні бактеріальним білкам RecA, RecBCD, RuvA, B і C. Наприклад, білок RAD51 дріжджів і людини є гомологічним білку RecA й виконує подібні функції. Ініціюючий рекомбінацію дволанцюговий розріз індукується у дріжджів білком Spo11, що належить до родини ДНК-топоізомераз ІІ. Гомологічні білки знайдено в інших еукаріотів і бактерій. Отже, механізми гомологічної рекомбінації є, імовірно, спільними для всіх біологічних систем.
Дата добавления: 2015-09-11; просмотров: 1850;