Глава 1. МОРФОЛОГИЯ И КЛАССИФИКАЦИЯ МИКРООРГАНИЗМОВ
Морфология микроорганизмов изучает форму и особенности строения клеток, способность двигаться, образовывать споры, способы размножения и др. По современным .представлениям все живые организмы, имеющие клеточное строение, делятся на два надцарства: прокариоты и эукариоты (греч. «ка-рион» —ядро). Организмы, не имеющие клеточного строения, составляют третье надцарство — акариоты (например, вирусы). К прокариотам относится только одно царство — бактерии, в том числе цианобактерии (сине-зеленые водоросли). К эука-риотам относятся три царства: животные, растения и грибы.
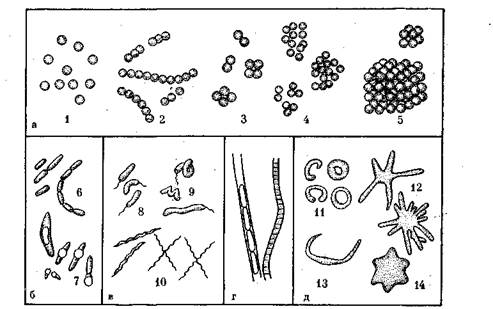
Рис. 1. Формы бактерий:
а — шаровидные; б — палочковидные; в — извитые; 4— нитчатые; д — новые формы- 1 — микрококки; 2 —стрептококки; 3 —диплококки и тетракокки;.. 4 — стафилококки; 5 —сардины; б —палочки без спор; 7 — палочки со спорами; 8"—вибрионы; 9 — спириллы; 10 — спирохеты; //— тороиды; 12 — бактерии, образующие простеки; 13 — червеобразные'; 14 — шестиугольные
Деление живых организмов на прокариоты и эукариоты базируется прежде всего на особенностях строения их ядерного аппарата. С помощью электронного микроскопа было установлено отсутствие у бактерий истинного ядра, поэтому их назвали прокариотами, то есть «доядерными» организмами. Известно, что основой ядерного аппарата является дезоксирибонук-леиновая кислота (ДНК), молекула которой имеет вид двойной спирально закрученной нити. Ядерный аппарат прокариот включает молекулу ДНК в виде замкнутой в кольцо нити, расположенную непосредственно в цитоплазме. Ядерный аппарат прокариот называют нуклеоидом, что в переводе с латинского означает «подобный ядру». У эукариот имеется истинное ядро с ядрышком, окруженное ядерной мембраной. Внутри ядра заключена ДНК. Наряду с этим основным признаком имеется много специфических особенностей в строении и обмене веществ прокариот.
Основными объектами технической микробиологии являются бактерии, мицелиальные грибы ,и дрожжи, которые в основном составляют как полезную, так и нежелательную микрофлору пищевых производств.
ПРОКАРИОТЫ (БАКТЕРИИ]
В мире микроорганизмов бактерии по численности (около 4000 видов) и разнообразию осуществляемых ими химических превращений занимают ведущее место. Большинство бактерий —это одноклеточные организмы, но имеются и многоклеточные.
Форма и размеры бактерий. Одноклеточные бактерии по внешнему виду разделяются на три основные группы: шаровидные, палочковидные и извитые (рис. 1).
Шаровидные бактерии —кокки (рис. \,а) могут быть одиночными — микрококки либо соединенными попарно — диплококки. Часто при делении клетки по тем или иным причинам не расходятся и образуют различные сочетания, которые зависят от расположения делящей перегородки. Когда делящие перегородки располагаются в двух взаимно перпендикулярных плоскостях, то образуются группы, состоящие из четырех клеток — тетракокки. При делении в трех взаимно перпендикулярных плоскостях образуются пакетообразные скопления, состоящие из восьми — шестнадцати кокков, называемые сарцинами. При делении кокков в разных направлениях образуются скопления клеток, напоминающие грозди винограда, — стафилококки. Если деление кокков происходит в одном направлении и они при этом не разделяются, то образуются цепочки клеток— стрептококки. Эти сочетания не эквивалентны многоклеточным микроорганизмам, так как каждая клетка в них является отдельным организмом, способным на самостоятельное существование после отделения от остальных клеток.
|
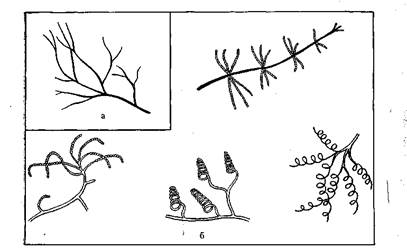
Рис. 2. Актиномицеты:
а — мицелий; б — спороносны
Палочковидные бактерии (рис. 1, б) имеют форму вытянутого цилиндра, могут быть одиночными или соединенными попарно, а также в виде цепочек из трех и более клеток. Отношение длины клетки к ее поперечнику у них сильно варьирует. У коротких палочек длина лишь ненамного превышает поперечное сечение и их иногда довольно трудно отличить от кокков. Палочковидные бактерии являются наиболее многочисленной группой среди бактерий.
Извитые (рис. 1, в) бактерии бывают трех типов: вибрионы— палочки, изогнутые в виде запятой; спириллы, имеющие несколько правильных завитков, и спирохеты, имеющие вид мелких спиралей с многочисленными завитками.
Кроме этих наиболее распространенных в природе форм бактерий имеется небольшое число нитчатых форм (рис. 1, г). Они представляют собой многоклеточные организмы в виде ,нитей, состоящих из одинаковых цилиндрических или дисковидных клеток.
Сравнительно недавно в почве и водоемах были обнаружены новые формы бактерий, клетки которых имеют вид разомкнутого или замкнутого кольца (тороиды), шестиугольной звезды, розетки, а также клетки с выростами (просте-ками) и червеобразной формы (рис. 1, д).
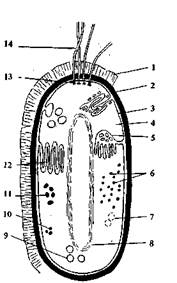
Рис. 3. Схема строения бактериальной клетки: 1 — капсула; 2 — клеточная стенка; 3 — цитоплазмати-ческая мембрана; 4— цитоплазма; 5 — мезосомы; 6 — рибосомы; 7 — полисахаридные гранулы; 8 — нуклео-ид; 9 — включения серы; 10 — жировые капли; 11 — полифосфатные гранулы; 12 — внутриплазматические мембранные образования; 13 — базальное тельце; 14 — жгутики
К бактериям относится еще одна, особая группа микроорганизмов — актиномицеты. Их клетки в основном имеют вид очень тонких длинных прямых ветвящихся нитей (рис. 2).
Размеры бактерий ничтожно малы, поперечное сечение клеток большинства бактерий не превышает 0,5—0,8 мкм, средняя длина палочковидных бактерий от 0,5 до 3 мкм. Нитчатые бактерии значительно крупнее — не-
которые имеют 15-—125 мкм в длину и 5—35 мкм в поперечнике. Длина клеток спирохет может достигать 500 мкм. Наиболее мелкие из микроорганизмов — микоплазмы, не имеющие клеточной стенки, имеют размер 0,1—0,15 мкм.
Объем бактериальной клетки в среднем составляет 0,07 мкм3, масса — 5-10~12 г. В 1 мм3 может содержаться до 109 бактериальных клеток.
В пищевых производствах основное значение имеют шаровидные и палочковидные бактерии.
Строение, химический состав и функции клеточных структур бактериальной (прокариотной) клетки. Обязательными клеточными структурами у преобладающего большинства бактерий являются: клеточная стенка, цитоплазматическая мембрана (ЦП*М), ядерный аппарат (нуклеоид) и рибосомы (рис.3).
Клетка снаружи покрыта жесткой клеточной стенкой. Она придает форму клетке, предохраняет ее от неблагоприятных внешних тепловых и механических воздействий, защищает клетку от проникновения в нее избытка воды. У некоторых бактерий на наружной .поверхности клеточной стенки образуются капсулы или слизистый слой. Капсула чаще всего состоит из полисахаридов (декстрана, левана), реже —из полипеп-тидов. Капсула — необязательная структура бактериальной клетки. Иногда капсулы служат источником запасных пита-тательных веществ. Например, капсулы из полисахаридов образуются у клеток лейконостока на средах с значительным количеством углеводов.
По химическому составу и строению клеточной стенки бактерии делятся на 2 большие группы: грамположительные и грамотрицательные бактерии (Трам+ и Грам-).
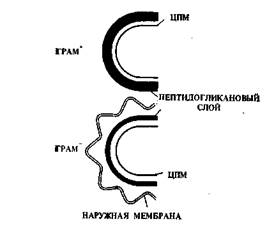
гис. *. ч.лема строенияклеточных стенок грамположительных и грамотрицательных бактерий
Названы так по имени датского ученого Кристиана Грама, предложившего специальный способ окраски бактерий (окраска по Граму). После окраски препарат бактерий обрабатывают спиртом или ацетоном, в результате чего Грам- бактерии обесцвечиваются, а Грам+ бактерии сохраняют темно-фиолетовую окраску. Окраска по Граму имеет важное значение для классификации бактерий.
Как у Грам+, так и у Грам- бактерий жесткость
- клетОЧНОЙ СТеНКИ Обусловлена наличием полимерного соеди-
нения пептидогликана (муреи-на), но у Грам+ бактерий его количество значительно больше (до 90—95% от веществ клеточной стенки), а у Грам-— 5—10%. Пептидогликановый слой у Грам+ бактерий плотно прилегает к ЦПМ (рис. 4).
Кроме того, в клеточных стенках Грам+ бактерий имеются другие полимеры — тейхоевые кислоты, которые так же, как и пептидогликан, имеются только у прокариот, а у эукариот не обнаруживаются. В составе клеточной стенки Грам+ бактерий в небольших количествах содержатся полисахариды. У Грам+ бактерий клеточная стенка имеет толщину 20—80 нм, юна однослойная и плотная.
Клеточная стенка Грам- бактерий значительно тоньше — 10—13 нм, но она многослойная. Пептидогликан образует только внутренний слой, неплотно прилегающий к ЦПМ. К внутреннему слою прилегает наружная мембрана, состоящая из липопротеидов и липополисахаридов. Тейхоевые кислоты в клеточной стенке Грам- бактерий отсутствуют.
Наружная мембрана Грам- бактерий препятствует проникновению в клетку токсических веществ, поэтому Грам- бактерии значительно устойчивее по сравнению с Грам+ бактериями к действию антибиотиков, ядовитых химических и др. веществ. Поэтому в пищевых производствах борьба с Грам- бактериями с помощью дезинфицирующих средств не всегда эффективна.
Цитоплазматическая мембрана (ЦПМ) расположена под клеточной стенкой, ограничивает содержимое клетки и играет очень важную роль в жизни клетки. Нарушение ее целостности приводит к гибели клетки. Химически ЦПМ представляет собой белково-липидный комплекс, состоящий из белков (50— 75% от массы ЦПМ), липидов (в основном фосфолипиды — 15—45%) и небольшого количества углеводов. В ЦПМ имеются поры, через которые в клетку поступают питательные вещества и выводятся конечные продукты обмена веществ.
Поскольку у прокариот ЦПМ единственная, в отличие от эукариот, мембранная структура в клетке, то она выполняет много функций: осуществляет транспорт питательных веществ из внешней среды внутрь * клетки с помощью специфических белков — переносчиков; на внутренней стороне ЦПМ расположены окислительно-восстановительные ферменты, участвующие в снабжении клетки энергией, и гидролитические ферменты, рсуществляющие расщепление высокомолекулярных соединений. У некоторых бактерий ЦПМ образует впячивания внутрь клетки — мезосомы, имеющие различные формы и размеры и выполняющие различные функции (участие в энергетических процессах, в процессах деления клетки, процессе размножения и др.).
Цитоплазма — это внутреннее содержимое клетки, окруженное ЦПМ, представляющая собой полужидкую коллоидную систему. Она содержит воду до 70—80% от массы клетки, ферменты, аминокислоты, набор РНК, субстраты и продукты обмена веществ клетки. В цитоплазме располагаются остальные жизненно важные структуры клетки — нуклеоид, рибосомы, а также запасные вещества различной природы.
Нуклеоид представляет собой ядерный аппарат прокариот. Это компактное образование, занимающее центральную область в цитоплазме, состоящее из двойной спирально закрученной нити ДНК, замкнутой в кольцо, которая еще называется бактериальной хромосомой. Бактериальная хромосома в одной точке соприкасается с мезосомой. В развернутом виде нить ДНК может иметь длину более 1 мм, т. е. почти в 1000 раз больше длины бактериальной клетки. Вся генетическая информация у прокариот, так же как и у эукариот, заключена -в ДНК, поэтому функция нуклеоида состоит в передаче наследственных свойств. Перед, делением клетки нуклеоид делится пополам. Ядерный аппарат прокариот не имеет ядрышка и не отделен от цитоплазмы ядерной мембраной, как это имеет место у эукариот.
Рибосомы — небольшие гранулы, рассеянные в цитоплазме, состоящие из РНК (60%) и белка (40%). Они играют очень важную физиологическую роль, поскольку на них происходит синтез белков. В молодых клетках наблюдается повышенное содержание рибосом.
В клетках бактерий, помимо обязательных клеточных структур, имеются включения запасных веществ. Они накапливаются при избытке тех или иных питательных веществ в среде, а расходуются при голодании клетки. К запасным веществам клетки бактерий относятся полисахариды, включающие гликоген, крахмал и гранулезу; жировые капли, содержащие липиды (жиры) в виде поли-р-оксимасляной кислоты, которая синтезируется на средах богатых углеводами. Поли-р-оксимасляная кислота встречается только у прокариот и ее количество может достигать 50% от сухой массы клеток. Гранулеза и липиды служат хорошим источником углерода и энергии для клетки. У многих прокариот в клетках накапливаются полифосфаты в виде гранул, называемых также валютиновыми или мета-хроматиновыми зернами. Они используются клетками как источник фосфора.
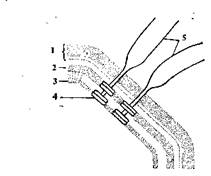
Рис. 5. Схема прикрепления
жгутика:
1 — клеточная стенка; 2 — цито-
плазматическая мембрана; 3 —
мембрана жгутиков; 4—диски
основания; 5 — жгутики
В клетках некоторых бактерий, участвующих в превращениях серы,, откладывается молекулярная сера в виде особых включений.
Подвижность бактерий.Способно-
стью к движению обладает примерно Уб часть бактерий. Это в основном многие палочковидные и все извитые формы бактерий. Неподвижными являются почти все шаровидные бактерии (кокки), более 50% палочковидных бактерий и ряд других.
Чаще всего движение осуществляется с помощью жгутиков (см. рис. 3) —тонких нитей толщиной 10—20 нм, состоящих из особого белка флагеллина. Длина жгутиков во много раз может превышать длину клетки. Жгутики (рис. 5.) прикрепляются к мембране с помощью двух пар дисков основания и через поры в ней и клеточной стенке выходят наружу. Скорость перемещения бактерий спомощью жгутиков высока (20— 60 мкм/с).
Характер расположения жгутиков на поверхности клетки является одним из признаков классификации бактерий (рис. 6). Их количество может быть от 1 до 100. Бактерии, имеющие один жгутик на конце клетки, называют монотрихами; с пучком жгутиков на одном или обоих концах клетки — лофотри* хами; один жгутик на обоих донцах — амфитрихами. Бактерии, у которых жгутики покрывают всю поверхность клетки,, называются перитрихами. Жгутики обеспечивают активное движение клеток только в жидкой среде, и при утрате жгутиков при старении или механическом воздействии клетки теряют способность к движению, но сохраняют жизнеспособность.
К подвижным формам относятся также спирохеты, некоторые нитчатые (многоклеточные) и другие бактерии, не имеющие . жгутиков. Спирохеты могут передвигаться и в жидкой среде, и по твердому субстрату в результате волнообразных сокращений клетки. Нитчатые бактерии, цианобактерии и другие обладают скользящим типом движения по твердому и полутвердому субстрату.
Способность к движению позволяет бактериям переместиться в ту область среды, в которой условия для их роста и размножения (концентрация питательных веществ и кислорода в среде, освещенность и др.) наиболее оптимальны.
Рис. 6. Расположение жгутиков у подвижных форм бактерий: а — монотрих; б — амфитрих; в — лофотрих; г — перитрих
Рост и размножение бактерий.Основной отличительной особенностью живых организмов от неживой природы является , рост и размножение. Рост — это физиологический процесс, в ходе которого увеличиваются размеры и масса клетки. Рост •бактериальной клетки ограничен, и, достигнув определенной величины, она перестает расти. Начинается процесс размножения, т. е. увеличение числа особей (клеток), когда от материнской клетки отделяется дочерняя.
Большинство бактерий размножается простым делением на две части. Такой способ размножения называется бинарным поперечным делением. У подавляющего большинства Грам+ бактерий клетки делятся ровно пополам с помощью септы (поперечной перегородки). На противоположных сторонах внутренней части клеточной стенки образуются два выступа, растущие навстречу друг другу (от периферии к центру), в этих же местах ЦПМ образует мезосомы (впячивания). Расположенные в мезосомах ферменты синтезируют материал клеточной стенки. Поперечная перегородка первоначально фомиру-€тся из ЦПМ и пептидогликана; наружные слои синтезируются позднее.
Клетки большинства Грам- бактерий делятся путем образования перетяжки. В центре клетки с одной стороны ЦПМ иклеточная стенка постепенно прогибаются до слияния с противоположной поверхностью клетки. Образованию поперечной перегородки или перетяжки предшествует деление ДНК, в результате чего в каждую дочернюю клетку попадает по одному нуклеоиду.
Актиномицеты размножаются главным образом экзоспора-ми (наружными спорами), которые образуются поодиночке или цепочками на концах спороносящих гиф— спороносцах, имеющих самую разнообразную форму (см. рис. 2). Существуют и другие способы размножения.
Образование эндоспор.Способностью к образованию эндоспор (внутренних спор) обладают лишь некоторые палочковидные Грам+ бактерии. Поскольку в каждой клетке образуется только одна спора, то спорообразование является не спо-
 |

|
|
а • «
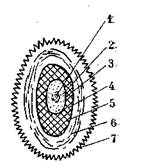
Рис. 7. Типы спорообразования у бактерий:
а — бациллярный; б — клостридиальный; в — плектридиальный
собом размножения, а покоящейся стадией клетки для перенесения неблагоприятных условий. Споры образуются при голодании, при избытке продуктов обмена веществ или несоответствии температуры, влажности и рН их оптимальным значениям для развития данного вида бактерий.
Различают три типа спорообразования (рис. 7). Если при образовании споры в центре клетки форма ее не меняется, то такой тип спорообразования называется бациллярным; он свойствен представителям рода ВасШиз. Если же клетка в середине утолщается и приобретает вид веретена, то такой тип спорообразования называется клостридиальным. Иногда спора образуется ближе к концу клетки и тогда клетка приобретает вид теннисной ракетки — такой тип спорообразования называется плектридиальным (рис. 7). Клостридиальный и плектридиальный типы спорообразования свойственны бактериям рода С1оз1:пс1шт.
Спорообразование — сложный процесс, в результате которого в клетке формируется эндоспора, отличающаяся от вегетативной клетки структурой и химическим составом (рис. 8). Эндоспора имеет наружную и внутреннюю мембраны, между которыми располагается кортекс (кора), сходный по химическому составу с клеточной стенкой вегетативной клетки. Поверх наружной мембраны образуются многослойные покровы споры, состоящие в основном из белков. У некоторых бактерий снаружи споры формируется еще один слой — экзоспориум, состоящий из липидов и белков.
При спорообразовании происходит накопление специфического вещества — дипиколиновой кислоты, отсутствующей в вегетативной клетке, а также ионов кальция. Процесс образования споры протекает * несколько часов. Когда спора сформируется, оболочка и остальные части клетки разрушаются и спора освобождается.
Рис. 8. Схема строения бактериальной споры:
/ — нуклеоид; 2 — цитоплазма; 3 — внутренняя мембрана; 4 — кортекс; 5 — наружная мембрана; 6 — покровы, состоящие из нескольких слоев; 7 — экзоспориум
Споры необычайно устойчивы к воздействию температуры, например, споры возбудителя тяжелого пищевого отравления — ботулизма — выдерживают нагревание до 100 °С в течение 5—6 ч. Споры выносят высушивание, воздействие ультрафиолетовых лучей, ядовитых веществ и т. п. Устойчивость спор связана с тем, что их покровы труднопроницаемы, в них содержите» много липидов, а также дипиколиновой кислоты и кальция. Активность ферментов в них подавлена. Высокая термоустойчивость спор обусловливается низким содержанием в них воды,, что предохраняет белки от денатурации при высоких температурах.
Споры бактерий могут сохранять жизнеспособность десятки и даже сотни лет. Попав в благоприятные условия, спора поглощает воду и набухает, ее термоустойчивость снижается, возрастает активность ферментов, под действием которых растворяются оболочки, и спора прорастает в вегетативную» клетку.
Порчу пищевых продуктов вызывают лишь вегетативные клетки бактерий. Поэтому необходимо знать условия, способствующие образованию спор и их прорастанию в вегетативные клетки, чтобы правильно выбрать способ обработки пищевых продуктов с целью предотвращения их порчи под влиянием бактерий.
Принципы классификации бактерий.В настоящее время типичной классификации бактерий нет, хотя работа по ее созданию ведется постоянно. Классификация всех живых существ основана почти полностью на непосредственно наблюдаемых и легко определяемых морфологических признаках организмов .У бактерий вследствие немногочисленности их морфологических признаков создать общепринятую классификацию невозможно и требуются дополнительные признаки.
Кроме того, организмы в соответствии с основными принципами классификации живых существ должны быть расположены в ряды от наиболее простых к более сложным, т. е. как шло их постепенное развитие (эволюция). Такая классификация организмов является естественной. Самой мелкой единицей классификации является вид— группа организмов, наделенная общими стабильными признаками и происходящая от общего предка. Близкородственные виды группируются в более высокую систематическую единицу — род; близкие роды — в семейства, семейства — в порядки или отряды, порядки — в классы, а классы — в типы.
Однако микробиологи в настоящее время не располагают достаточными знаниями об эволюции бактерий. Поэтому большинство имеющихся классификаций бактерий являются искусственными. Искусственные классификации предназначаются для определения той или иной группы микроорганизмов, которая .представляет практический интерес для исследователя.
Научные названия микроорганизмов состоят из двух латинских слов: первое пишется с прописной буквы и означает род, второе — со строчной буквы и означает вид данного рода. Например: ВасШиз зиМШз (палочка сенная) — это бактерия, относящаяся к роду ВасШиз, палочковидной формы, образующая эндоспоры бациллярного типа, постоянно обитает на сене.
Для классификации бактерий используют в основном следующие признаки: морфологические (форма клеток, наличие и характер расположения жгутиков, способ размножения, окраска по Граму, наличие эндоспор); физиологические (отношение к воздействию температуры, рН, кислорода, тип питания, способ получения энергии, характер образующихся .продуктов); культуралъные (характер роста на различных питательных средах культуры бактерий в массе, а не в виде отдельных клеток: на жидких средах это наличие пленки, мути, •осадка; на плотных средах — тип колоний и их особенности).
В последние годы получила признание классификация бактерий, предложенная Р. Мюрреем в 1978 г. Это искусственная классификация, в основу которой положено строение клеточной стенки. Все бактерии, для которых характерно строение клеточной стенки по типу Грам+ бактерий, отнесены в отдел Тчгтаси1.ез*. Другой отдел — ОгасШси1ез — объединяет ,все •бактерии, которые имеют клеточную стенку, характерную для Грам- бактерий. Третий отдел объединяет особые формы бактерий, лишенные настоящей клеточной стенки; они не играют роли в пищевых производствах и поэтому рассматриваться не •будут. Бактерии, имеющие значение в пищевых производствах, •относятся к первым двум отделам.
Отдел р1гтаси1ез. К нему относятся 4 группы; в основу деления на группы положена форма клеток и способность образовывать эндоспоры и экзоспоры. Это кокки, две группы палочковидных бактерий, актиномицеты и родственные организмы.
Кокки характеризуются округлой формой; деление клеток происходит в одной или нескольких плоскостях, при этом образуются различные сочетания клеток; кокки неподвижны, не образуют эндоспор. Многие микрококки являются возбудителями порчи пищевых продуктов, лейконосток является вредителем в сахарном производстве; некоторые стафилококки, развиваясь в пищевых продуктах, вырабатывают ядовитые ве-
* От лат. «кутикула» — кожица, «фирма» — солидная, «гратия» — изящная.
щества (токсины) и вызывают пищевые отравления. Сюда же* относятся молочнокислые стрептококки, используемые при производстве кисломолочных продуктов, маргарина, сливочного масла и др.
Вторая группа — это палочки, образующие эндоспоры. К ним относится одно семейство, представители которого очень широко распространены в природе. Это одиночные илц соединенные в цепочки палочки, многие из них подвижны,, имеют перитрихиальное жгутикование. Палочки образуют эндоспоры бациллярного типа (род ВасШиз) и клостридиаль-ного или плектридиального типа (род СЛозхгМшт). Многие являются .возбудителями порчи пищевых продуктов (например, гнилостные, маслянокислые бактерии). Имеется много возбудителей инфекционных болезней (сибирской язвы, столбняка) и пищевого отравления — ботулизма.
Третья группа — это палочки, не образующие эндоспор. К ним относится только одно семейство, включающее род Ьас1оЪасШиз. Это палочковидные, не образующие спор молочнокислые бактерии*. Чаще они представляют собой одиночные длинные и тонкие палочки, иногда — короткие палочки цепочками. Являются вредителями в процессах бродильных производств. Используются в производстве кисломолочных продуктов, в .сыроделии, квашении овощей, в хлебопечении.
Четвертая группа — актиномицеты и родственные организмы. Актиномицеты — своеобразная группа бактерий, представляющих собой длинные тонкие ветвящиеся нити без перегородок, называемые гифами, переплетение которых образует мицелий. Нижняя часть мицелия, врастающая в субстрат, называется субстратным мицелием и служит для обеспечения организма питанием, верхняя часть мицелия возвышается над субстратом и называется воздушным мицелием. Размножаются актиномицеты экзоспорами, образующимися в спороносцах. Некоторые из актиномицетов представляют собой короткие ветвящиеся палочки. Встречаются на пищевых продуктах, могут вызвать их порчу, при которой продукты приобретают отчетливый землистый запах. Имеются и патогенные виды (туберкулезная и дифтерийная палочки). Актиномицеты являются основными продуцентами антибиотиков, получаемых в промышленных масштабах, а также витаминов группы В (Вь В2, В3, В6, В12).
Отдел (Зг асШсихез. Все представители Грам^ бактерий не образуют спор и резко различаются по способности развиваться на свету и без него. Бактерии, встречающиеся в пищевых производствах, безразличны к свету. Они различаются по форме клеток и способу движения. По числу предста-
* Несмотря на то, что представители этого рода — палочки, не образующие спор, в научной литературе за ними сохраняется старое название Ьас1о-ЪасШиз.
|
вителей и значимости в природе и жизни человека наибольший интерес из них представляют псевдомонады и энтеробак-терии.
Из псевдомонад для пищевых производств наибольшее значение имеет обширный род Рзеиёотопаз. Это одиночные подвижные палочки с одним или с пучком полярных жгутиков (монотрихи и лофотрихи). Псевдомонады очень широко распространены в природе, активно участвуют в круговороте веществ, часто обнаруживаются в водоемах и почве, загрязненных различными соединениями, например пестицидами, участвуют и в их разложении. Многие из псевдомонад образуют флюоресцирующие пигменты, выделяющиеся в среду, и вызывают порчу пищевых продуктов (некоторые гнилостные, жиро-окисляющие и другие бактерии).
К Грам- палочкам относятся также уксуснокислые бактерии родов Асе1оЪае{ег (перитрихи) и 01исопоЬас1ег (моно-трихи), используемые в производстве уксуса. Многие из них являются вредителями в бродильных производствах.
В пищевых производствах наибольшее значение имеет многочисленная кишечная группа бактерий — энтеробактерии. Это одиночные подвижные палочки, перитрихи, но встречаются и неподвижные формы. Некоторые из них постоянно населяют кишечник человека и животных (например, кишечная палочка), другие являются возбудителями инфекционных желудочно-кишечных заболеваний (дизентерии, брюшного тифа, паратифов), передающихся через пищевые продукты, а также возбудителями пищевых отравлений.
Классификация бактерий, имеющих значение в пищевых производствах и рассматриваемых в данном курсе, приведена на с. 20.
ЭУКАРИОТЫ (МИЦЕЛИАЛЬНЫЕ ГРИБЫ И ДРОЖЖИ)
Одним из трех царств, относящихся к надцарству ,эукариот, являются грибы. Ранее считали, что грибы занимают промежуточное .положение между царствами растений и животных, так как ряд признаков сближает их как с животными, так и с растениями. Но в настоящее время грибы выделены в отдельное царство Мусо1а. Эта обширная и разнообразная группа организмов включает до 100 тыс. видой.
Грибы широко распространены в природе. Они обитают в различных климатических зонах от тропиков до Арктики, •особенно много их в почвах, в том числе высокогорных, на растениях; встречаются они в пресных и соленых водоемах, в местах с повышенной влажностью и т. п. Грибы для своего развития нуждаются в органических веществах.
Среди грибов встречаются организмы, развивающиеся за счет органических веществ отмерших организмов; они участвуют в круговороте веществ в природе. Но имеются и такие,
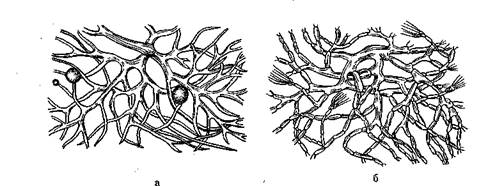
Рис. 9. Мицелий грибов:
а — несептированный; б — септированный
которые могут существовать только в живых организмах и вызывать их заболевания. Некоторые из грибов выделяют ядовитые вещества — микотоксины. Многие грибы вызывают порчу пищевых продуктов.и повреждение разнообразных изделий и материалов, некоторые могут развиваться даже на оптических поверхностях, где имеется мизерное количество смазки. Они утилизируют смазку и вызывают помутнение линз. Но грибы имеют и важное практическое значение, многие из них употребляются в пищу, используются в производстве этилового спирта, органических кислот, ферментов, антибиотиков, витаминов, некоторых сортов сыра и т. д.
Мицелиальные грибы. Царство грибов делится на семь классов, но объектами изучения микробиологии являются в основном три, включающие мицелиальные грибы, — зигомицеты (ранее их называли плесневые грибы), аскомицеты и дейтеро-
мицеты.
Форма и размеры. Клетки мицелиальных грибов имеют вытянутую форму в виде нитей (гифов), размеры которых достигают до 5—30 мкм в диаметре, что значительно превышает размеры бактериальной клетки.
Переплетение гиф образует тело гриба — мицелий, или грибницу (рис. 9). Большая часть гиф развивается над поверхностью субстрата (воздушный мицелий), на которой располагаются органы размножения, а часть — в толще субстрата (субстратный мицелий). Гифы у большинства мицелиальных грибов многоклеточные, в их клетках имеются поперечные перегородки — септы. Такой мицелий называют септирован-ным, он имеется у аскомицетов и дейтеромицетов. Мицелий зигомицетов несептированный и представляет собой одну гигантскую клетку с несколькими ядрами. Гифы растут за счет верхушечных клеток, и клетки гиф неодинаковы по длине.
Некоторые грибы на определенной стадии развития образуют плодовые тела, внутри которых находятся органы раз-
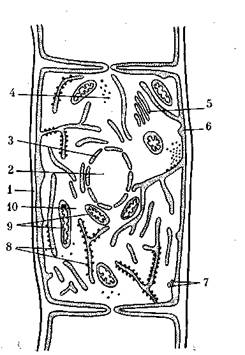
Рис. 10. Схема строения грибной
клетки:
1 — клеточная стенка; 2 — ядро; 3 — ядерная мембрана; 4 — рибосомы; 5 — аппарат Гольджи; 6 — цитоплазматиче^кая мембра-»на; 7 —лизосомы; 8 — эндоплазматическая сеть; 9 — митохондрии; 10 — цитоплазма
множения, покрытые сверху плотным переплетением гиф. У других видов грибов из плотных переплетений сильно разветвленных гиф образуются склероцш, богатые запасными питательными веществами. Они служат для перенесения неблагоприятных условий и являются покоящейся формой гриба.
Мицелиальные грибы не имеют жгутиков и относятся к неподвижным организмам.
Строение клетки. У мицелиальных грибов клетки
имеют строение, характерное
для клеток эукариотных микроорганизмов (рис. 10). У них имеется хорошо развитая система внутриклеточных элементарных биологических мембран (в отличие от прокариот, у которых только одна мембранная структура внутри клеток — ци-топлазматическая мембрана). Внутриклеточные структуры эу-кариот, полностью'ограниченные от цитоплазмы такими мембранами, называются органеллами. К органеллам кроме ЦПМ .относятся ядро, митохондрии, эндоплазматическая сеть, аппарат Гольджи и лизосомы.
Снаружи клетка мицелиальных грибов покрыта многослойной жесткой клеточной стенкой, состоящей на 80—90% из полисахаридов. Основным из них является азотсодержащий полисахарид хитин. Полисахариды связаны с белками, липи-дами, полифосфатами. Под клеточной стенкой расположена ЦПМ, которая окружает цитоплазму. В цитоплазме расположено ядро; оно содержит ядрышко, хромосомы и окружено ядерной мембраной с порами. В ядрышке синтезируются и скапливаются предшественники рибосом, которые затем транспортируются через поры ядра в цитоплазму. У грибов в длетках бывает от одного до 20—30 ядер. В цитоплазме рассеяны рибосомы.
Митохондрии — мембранные структуры, играющие очень важную роль. Они представляют собой многокамерные мешочки или трубочки с эластичными стенками, образующими впячивания — кристы (рис. 11). На них находятся окислительно- 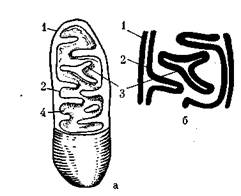
Рис. 11. Схема строения митохондрий:
а — общая схема строения; б —продольный разрез митохондрии; / — наружная мито-хондриальная мембрана; 2 — внутренняя митохондриальная мембрана; 3 — кристы; 4 — матрикс
восстановительные ферменты (у прокариот эти ферменты локализованы в ЦПМ), участвующие в энергетическом обмене. Поэтому митохондрии называют «силовыми станциями клетки», «энергетическими ансамблями» и т. п.
Эндоплазматическая сеть —-мембранная система, состоящая из канальцев, пузырьков или цистерн, которые не имеют строго определенной локализации, а располагаются либо по периферии клетки, либо вокруг ядра, либо пронизывают всю цито
плазму. На них расположены различные ферменты, ответственные за синтез липидов, углеводов, за транспорт веществ внутри клетки. Аппарат Гольджи — система мембран, связанных с ядерной мембраной и мембранами эндоплазматической сети. Он расположен на участке цитоплазмы, где нет рибосом. Роль аппарата Гольджи полностью не выяснена. Предполагают, что в аппарате Гольджи происходит синтез материала клеточной стенки и новых мембран, а также с его помощью осуществляется транспортирование веществ, синтезируемых в эндоплаз* матической сети, и удаление из клетки продуктов обмена.
Лизосомы представляют собой мембранные структуры округлой формы. В них сосредоточены гидролитические ферменты (у прокариот они локализованы в ЦПМ), расщепляющие белки, полисахариды, липиды.
В клетках мицелиальных грибов хорошо видны вакуоли — полости, окруженные мембраной и заполненные клеточным соком. Обычно они располагаются вблизи клеточной стенки, их число увеличивается по мере старения клеток. Основные запасные питательные вещества мицелиальных грибов — гликоген, который образуется на средах с избытком сахара; метахро-матин, который в виде гранул находится в самих вакуолях, а в цитоплазме вблизи вакуолей накапливаются липиды в виде жировых капель.
Размножение и классификация. Мицелиальные грибы размножаются бесполым и половым путем. Оба способа размножения связаны с образованием спор — наружных (экзо-споры) и внутренних (эндоспоры). Образованию спор при половом размножении предшествует процесс слияния содержимого двух клеток и их ядер. Вновь образовавшееся ядро делится на несколько частей — спор. Кроме того, все грибы
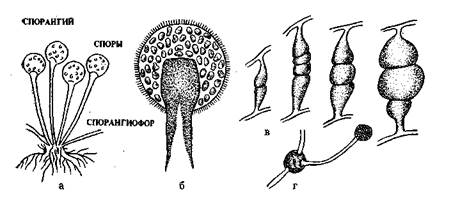
Рис. 12. Зигомицеты:
1 — КЬ12ориз; б — Мисог — спорангий с эндоспорами; в — последовательные стадии образования зигоспоры при половом размножении; г — проросшая зигоспора со спорангием
могут размножаться вегетативно — путем верхушечного роста
гиф, а также с помощью кусочков гиф и мицелия. Грибы, спо
собные к половому размножению, относятся к совершенным
(аскомицеты, зигомицеты), а те, которые не имеют полового
размножения, относятся к несовершенным грибам (дейтеро-
мицеты). У грибов имеется большое разнообразие способов
и органов размножения. \
Класс 2у^отусе1;е5 (зигомицеты). Это наиболее просто организованные грибы. Мицелий у них несептирован-ный, многоядерный, имеет вид одной гигантской разветвленной клетки. К зигомицетам относятся- мукоровые грибы. Они широко распространены в природе. Наибольшее значение имеют лредставители родов Мисог и КЫгориз.
Размножаются зигомицеты бесполым и половым путем (рис. 12). При бесполом размножении^в особых шаровидных вздутиях — спорангиях, формирующихся на концах длинных плодоносящих гиф — спорангиеносцах, образуются эндоспоры, называемые спорангиоспорами. Спорангиеносцы бывают одиночными (у грибов рода Мисог) или собранными в пучки с корневидными разрастаниями у основания — ризоидами (у грибов рода КЫгориз).
При половом размножении вначале происходит слияние двух многоядерных гиф мицелия, которые представляют собой обычно короткие образования с небольшим утолщением на концах. Затем происходит попарное слияние ядер. Заканчивается половое размножение образованием зиготы (зигоспоры), которая после периода покоя прорастает и образует короткую гифу со спорангием на конце. При прорастании споры происходит деление ядер. Многоядерная цитоплазма спорангия распадается на множество спорангиоспор, которые в благоприятных условиях могут прорасти чв мицелий.
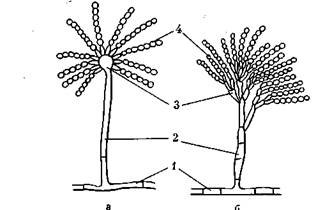
Рис. 13. Конидиеносцы аскомицетов: а — у грибов рода АзрегдШиз; б — у грибов рода РегпсПшт; / — вегетативный мицелий; 2 — конидие-носец; 3 — фиалиды; 4 — конидии
Многие грибы рода* Мисог вызывают порчу пищевых продуктову образуя пушистые серые налеты. Грибы рода КЫгориз вызывают так называемую «мягкую гниль» ягод, плодов и овощей. Муко-ровые грибы образуют органические кислоты и ферменты, способны вызывать слабое спир1-товое брожение, в связи с чем они применяются в некоторых странах Востока для получения напитков.
Класс Аз сот у-се1;е5 (а с коми це-ты, или сумчатые грибы). К ним относятся представители широко распространенных грибов родов РешсШшгп и Аз-рег^Шиз.
Аскомицеты имеют хорошо развитый многоклеточный мицелий. Бесполое размножение у них происходит с помощью зкзоспор, называемых конидиями, которые формируются на концах специализированных гиф — конидиеносцев. У аспергил-лов они простые, без перегородок, вздутые на вершине в виде пузыря, на котором располагаются фиалиды, отчленяющие цепочки шаровидных конидий. У пенициллов конидиеносцы многоклеточные, в виде кисточки, состоящей из мутовок фиа-лид (рис. 13). Конидии бывают различной окраски (зеленые, желтые, черные, голубые и т. п.). Конидии распространяются воздушными потоками, насекомыми, каплями росы, дождя и, прорастая, образуют новый мицелий.
Половое размножение аскомицетов происходит путем слияния содержимого и ядер двух клеток разных гиф, после чего происходит деление ядра; вокруг новых ядер концентрируется цитоплазма и образуется споровая оболочка. Материнская клетка покрывается толстой оболочкой и превращается в аск (сумку), внутри которого чаще всего находится 8 аскоспор. Сверху сумка покрывается переплетением гиф, образуя плодовое тело.
Большинство видов аспергиллов и пенициллов существуют только в конидиальной стадии и не имеют сумчатой стадии спороношения, поэтому такие виды отнесены к классу дейте-ромицетов (несовершенных грибов). На пищевых продуктах сумчатые грибы встречаются только в конидиальной стадии. Сумчатые грибы широко распространены в природе. Многие из них являются возбудителями порчи плодов и овощей, особенно при их хранении (различные гнили), а также многих пищевых продуктов, промышленных изделий и материалов (текстиля, резины, целлофана, пластмасс и т. д.), вызывая их повреждение. Некоторые аспергиллы являются патогенными для человека и животных, вызывают заболевание верхних дыхательных путей, слизистой рта, кожи (аспергиллез). Другие виды аспергиллов, а также гриб спорынья (паразит злаковых растений) выделяют ядовитые вещества, вызывающие пищевые отравления.
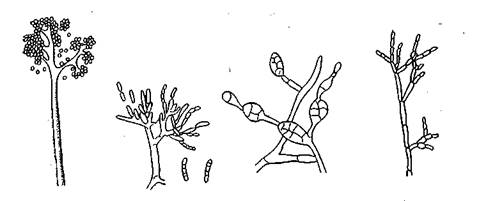
Рис. 14. Конидиеносцы и конидии различных родов несовершенных грибов: а — Во1гуИз; б — Ризагшт; в — АИегпаНа; г — С1ас1о5ропит
Однако некоторые представители сумчатых грибов нашли практическое применение. Так, отдельные представители пени-цилловых грибов используются как продуценты антибиотика пенициллина в промышленных масштабах, другие — в производстве сыра сортов «Рокфор», «Камамбер». Аспергиллы продуцируют органические кислоты, в связи с чем применяются для промышленного получения лимонной кислоты (Азрег§Шиз ш§ег). Многие аспергиллы используются для промышленного получения различных ферментных препаратов, используемых в отраслях пищевой и легкой промышленности.
Класс Оеи1еготусе1ез (дейтеромицеты). Дей-теромицеты, или несовершенные грибы, имеют многоклеточный мицелий. Половое размножение у них отсутствует, они размножаются только бесполым путем, в основном конидиями, которые, как и конидиеносцы, имеют самую различную форму и вид.
Конидиеносцы чаще всего многоклеточные, но могут быть одиночные — ветвящиеся или в виде пучков, со вздутиями. Конидии могут быть одноклеточными, многоклеточными, иногда с продольными и поперечными перегородками (рис. 14). По форме конидии бывают шаровидные, эллипсовидные, ните-
|
|
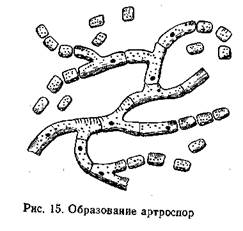
видные, серповидные, звездчатые и др. Некоторые дейтеро-мицеты (например, молочная плесень) размножаются не конидиями, а особыми клетками — артроспорами, которые образуются в результате фрагментации конидиеносца или гифы (рис. 15).
Несовершенные грибы широко распространены в природе. Большинство из них вызывает различные заболевания? растений и порчу продуктов. Так, представители рода Ри~ запит являются возбудителями заболевания плодов и овощей (фузариоз), вызывают порчу картофеля (сухая гниль). Некоторые виды этого гриба вырабатывают ядовитые для человека вещества, вызывающие тяжелые пищевые отравления. Грибы рода ВоггуИз вызывают порчу лука, капусты, моркови, помидоров, а вместе с другими грибами — кагатную гниль сахарной свеклы. Грибы рода А1-{егпапа поражают корнеплоды в период хранения (черная гчшль). Сердцевинную гниль свеклы вызывает гриб рода РЬо-та. Молочная плесень ОеоШсЬит сапсНйит вызывает порчу квашеных овощей, сметаны, творога и др., образуя на поверхности белую бархатистую пленку. Грибы из рода С1ас1озропига часто обнаруживаются на пищевых продуктах, хранящихся в холодильниках.
Единичные представители мицелиальных грибов включены в другие классы грибов. Это в основном грибы-паразиты, поражающие растения, используемые в качестве сырья для пищевых рроизводств. Например, гриб 5упспу{пшп — возбудитель рака клубней картофеля (класс СшЧгЫютусе^ез); гриб рода Рпук>рЫога поражает клубни и ботву картофеля, помидоры, и баклажаны (класс Оотусе!ез) и грибы рода головневых поражают зерновые культуры (например, пшеницу) и вызывают заболевание, называемое головней (класс Ваз1сПогпусе-1ез —высших грибов, куда относятся шляпочные грибы,, большинство из которых съедобно).
Дрожжи.Группа дрожжей объединяет одноклеточные грибные организмы, не имеющие настоящего мицелия.
Дрожжи широко распространены в природе. Они обитают в основном на растениях, где имеются сахаристые веще'ства, которые они сбраживают (нектар цветов, сочные фрукты, ягоды, особенно перезрелые и поврежденные, листья, стволы березы во время сокотечения и дуба во время слизетечения, почва). Переносятся дрожжи ветром, дождем и насекомыми.
Форма и размеры. Дрожжи могут иметь овальную, яйцевидную, округлую, лимоновидную, реже — цилиндрическую, треугольную, серповидную, стреловидную, колбо-видную формы клеток. Размеры дрожжей варьируют у разных видов от 1,5 — 2 до 10 мкм в поперечнике и до 2---20 мкм (иногда до 50 мкм) в длину.
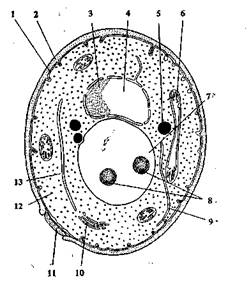
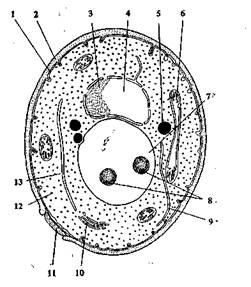
Рис. 1.6. Схема строения дрожжевой клетки:
1 — цитоплазматическая мембрана; 2 — клеточная стенка; 3— ядрышко; 4 — ядро; 5 — жировые капли; 6 — митохондрии; 7 — вакуоль; 8 — гранулы полифосфата; 9 — эндоплазматическая сеть; 10 — диктиосо-мы; 11 — почковый рубец; 12 — рибосомы; 13 — цитоплазма
.
Некоторые дрожжи на определенной стадии развития могут образовывать мицели-альные структуры — псевдомицелий. Дрожжи, как и все грибы, являются неподвижными организмами.
Строение клетки. Дрожжи, как и мицелиальные грибы,,, относятся к эукариотам и имеют сходное с ними строение-клетки, но имеются и некоторые различия (рис. 16). Клеточная стенка дрожжей, в отличие от грибов, на 60—70% состоит из полисахаридов глюкана и маннана, связанных с белками.' и липидами, и лишь небольшое количество (1—3%) составляет хитин, который вкраплен в стенку в виде гранул. У ряда? дрожжей в определенных условиях могут образовываться слизистые капсулы различной толщины полисахаридной природы.. Клетки таких дрожжей могут склеиваться друг с другом, образовывать хлопья и оседать на дно сосудов, в которых они развиваются.
Клетки дрожжей, как и грибов, имеют хорошо развитый:; мембранный аппарат — ЦПМ, эндоплазматическую сеть, аппарат Гольджи, лизосомы, митохондрии. В цитоплазме имеется-ядро. Рибосомы у дрожжей располагаются в цитоплазме и на? внешней стороне ядерной мембраны. Имеются вакуоли и включения запасных питательных веществ: липидов (особенно их много у дрожжей — продуцентов липидов), гликоген, метахро-матин. Клеточные структуры дрожжей выполняют те же-функции, что и у грибов.
Размножение и классификация. Дрожжи размножаются вегетативно и спорами, образующимися бесполым и половым путем. Способ размножения является важным-признаком для классификации дрожжей. К вегетативным способам размножения относятся: почкование, деление и почкующееся деление (рис. 17).
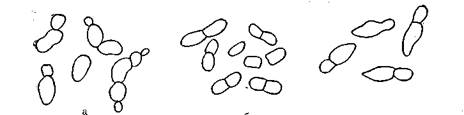
|
Способы вегетативного размножения дрожжей: почкование; а- почкование, б-деление; в - почкующееся деление
Почкование является .наиболее распространенным способом размножения дрожжей. При почковании на поверхности материнской (делящейся) клетки возникает маленький бугорок— почка, которая постепенно увеличивается дочти до размеров материнской клетки и превращается в дочернюю клетку. Она отделяется от материнской, оставляя на месте прикрепления почковый рубец. На этом месте почка больше не образуется. Может образовываться одна почка (полярное почкование), две почки на разных концах материнской клетки .(биполярное почкование), в нескольких местах поверхности материнской клетки (множественное почкование). Дочерние клетки могут не отделяться от материнской и оставаться соединенными с ней. Почкование характерно для дрожжей овальной и округлой формы.
У некоторых дрожжей при почковании дочерние клетки не отделяются от материнской, а вытягиваются в длину и продолжают образовывать все новые и новые почки, что приводит ж образованию ложного мицелия (псевдомицелий). Псевдоми-аделий характерен для пленчатых дрожжей.
Деление клетки в результате образования в ней поперечной перегородки — септы — характерно для дрожжей цилиндрической формы.
Почкующееся деление характеризуется тем, что образование дочерних клеток начинается с почкования, а заканчивается появлением хорошо заметной септы в районе перешейка. Такой способ размножения характерен для дрожжей лимоновидной формы.
Любому вегетативному способу размножения предшествует деление ядра, при котором одно из вновь образовавшихся ядер вместе с цитоплазмой и частью клеточных структур переходят в дочернюю клетку и они получают возможность самостоятельно существовать. У некоторых дрожжей имеется способ бесполого размножения с помощью бесполых спор, образующихся без слияния дрожжевых клеток. Бесполые споры — эндоспоры — нередко появляются в неопределенном ,числе в старых : культурах дрожжей, размножающихся делением и образующих мицелий.
Половое размножение у дрожжей происходит также с помощью спор, но их образованию предшествует процесс копуляции (слияние содержимого двух клеток и их ядер). Образуется зигота, в которой затем формируются споры: ядро делится, вокруг новых ядер уплотняется цитоплазма, • и они покрываются плотной оболочкой. Зигота со спорами внутри1 называется аском (сумкой), а споры— аскоспорами. Такие дрожжи относятся к классу аскомицетов, и их называют аскомицетовыми дрожжами. Аскоспоры могут образовывать только молодые клетки, выращенные на полноценной питательной среде и перенесенные в условия голодания, плохого снабжения кислородом и влагой. У различных видов дрожжей в аске образуется 2—4, а иногда 8 спор.
При благоприятных условиях аскоспоры выходят из аска и превращаются в вегетативные клетки. У некоторых видов дрожжей могут сливаться ядра материнской и дочерней клеток или ядра двух сестринских почек. Иногда происходит копуляция прорастающих спор соседних клеток.
Аскоспоры у дрожжей могут быть овальными, круглыми, бобовидными, игловидными, шлемовидными, кепковидными,. с гладкой, морщинистой поверхностью, с бородавчатыми или шиловидными выростами и т. д. Споры дрожжей, так же как и споры мицелиальных грибов, выполняют двойную функцию: служат для перенесения неблагоприятных условий, но главное,, в отличие от эндоспор бактерий, они служат для размножения. Споры дрожжей более устойчивы, чем вегетативные клетки, но* менее устойчивы, чем бактериальные споры. Так, споры дрожжей выдерживают нагревание при температуре на 10° больше,, чем вегетативная клетка (40—50 °С), а споры бактерий — на 50—60 °С больше, чем вегетативные клетки (60—120 °С).
Поскольку дрожжи по существу являются одноклеточными немицелиальными грибами, они включены в классификацию* грибов. Однако в отдельную систематическую единицу они не выделены, а распределены по трем классам грибов — аскомицетов, базидиомицетов и дейтеромицетов. Для микробиологии пищевых производств имеют значение лишь аскомицетовые и несовершенные дрожжи. Между этими дрожжами имеется принципиальное различие: у аскомицетовых дрожжей имеется. половой процесс и они вызывают энергичное спиртовое брожение. .Несовершенные дрожжи полового процесса не имеют и, как правило, вызывают слабое спиртовое брожение или вообще его не вызывают.
Аскомицетовые дрожжи. Включают примерно 2/3 дрожжей. Среди них наибольшее практическое значение имеют сахаромицеты, объединяющие более половины известных родов-дрожжей. Особо важная роль принадлежит роду сахаромицеты, все виды которого вызывают энергичное спиртовое брожение. Дрожжи этого рода размножаются бесполым способом (почкование) и с помощью аскоспор, которые образуют -половым путем.
В пищевых производствах наиболее широко используются -два вида дрожжей этого рода: Сахаромицес церевизия (крупные овальные клетки) в производстве этилового спирта, пива, -кваса и в хлебопечении и Сахаромицес эллипсойдес (крупные ^эллиптические клетки) — их используют преимущественно в виноделии. В каждом из этих производств применяют свои, •специфические расы (разновидности) данных видов дрожжей, обладающие наиболее ценными производственными свойствами.
К аскомицетовым дрожжам относятся и другие роды дрожжей. Это род Шизосахаромицеты, клетки которых имеют палочковидную форму и размножаются делением или с помощью «аскоспор, образующихся в результате полового размножения *(их количество 4—8). Дрожжи этого рода вызывают спиртовое брожение. Вид З Шизосахаромицес помбе используется в бродильной промышленности в странах с жарким климатом, например ,в Африке, где производят пиво сорта «Помбе». Дрожжи рода Сахаромикоды имеют крупные клетки лимоновидной формы. Они размножаются способом почкующегося деления на обоих концах 'клетки (биполярно) и с помощью аскоспор (их количество 2—4), которые расположены парами и образуются половым путем. Причем при половом способе происходит копуляция спор в аске, а не слияние зклеток дрожжей. Эти дрожжи вызывают спиртовое брожение, йю они являются вредителями в виноделии, так как образуют (продукты, придающие винам неприятный прокисший запах.
Некоторые аскомицетовые дрожжи используются в микробиологической промышленности для получения липидов и витаминов. Так, дрожжи рода Липомицес имеют крупные круглые клетки, которые в старых культурах заполнены целиком большой каплей жира. Обычно они имеют хорошо выраженные капсулы. Дрожжи рода Липомицес размножаются почкованием и аскоспорами, число которых у некоторых видов может доходить до 30 в одном аске.
Несовершенные дрожжи. Относятся к классу дей-теромицетов. Они не образуют спор, поэтому эти дрожжи част^, называют аспорогенными. Размножаются они почкованием. Несовершенные дрожжи вызывают либо слабое брожение, либо не вызывают его вообще, поэтому их часто называют несахаромицетами.
Многие из них являются причиной порчи пищевых продуктов и являются вредителями ряда пищевых производств. Однако некоторые из несовершенных дрожжей нашли полезное практическое применение. Среди несовершенных дрожжей наибольшее значение имеют роды Кандида, Торулопсис и Родоторула.
Дрожжи рода Кандида имеют удлиненную форму клеток, сочетания которых образуют примитивный псевдомицелий. Многие из них не вызывают спиртовое брожение и являются вредителями в бродильных производствах (например, Кандида микодерма), так как, будучи аэробами, окисляют спирт до ди: оксида углерода (углекислого газа) и воды. Другие представители рода Кандида являются вредителями в дрожжевом производстве, снижают качество хлебопекарных дрожжей, так как относятся к слабосбраживающим видам. Некоторые из них вызывают порчу квашеных овощей, безалкогольных напитков и ряда других продуктов. Среди этих дрожжей имеются патогенные виды, вызывающие кандидозы, поражающие слизистые оболочки ротовой полости, носоглотки и других органов человека. Различные виды дрожжей рода Кандида используются для получения кормового белка и белково-витаминных концентратов (БВК).
Дрожжи рода Торулопсис имеют мелкие круглые или овальные клетки. Многие виды способны вызывать слабое спиртовое брожение и используются в производстве кефира и кумыса. Некоторые применяются для промышленного получения кормового белка.
Дрожжи рода Родоторула имеют круглые, овальные или удлиненные клетки, последние образуют псевдомицелий. Колонии таких дрожжей красные и желтые благодаря наличию пигментов каротиноидов, являющихся провитамином А. Эти дрожжи используются для промышленного получения кормовых белково-каротиноидных концентратов, которые служат источником жирорастворимого витамина А для животных. Другие представители этого рода накапливают в клетках много липидов и используются в микробиологической промышленности как продуценты липидов наряду с представителями несовершенных дрожжей другого рода — Криптококкус.
.
ВИРУСЫ
Изобретение электронного микроскопа позволило впервые наблюдать мельчайшие организмы — вирусы и фаги. Вирусы часто называют фильтрующимися за их способность проходить через поры бактериологических фильтров, которые задерживают бактерии при механическом способе стерилизации. Вирусы были открыты в 1892 г. русским ботаником Д. И. Ивановским при изучении болезни табака — табачной мозаики. Их размеры колеблются от 10—12 нм (вирусы ящура, полиомиелита) до 200—350 нм (вирусы оспы, герпеса).
Вирусы не имеют клеточного строения. Они бывают шарообразной, палочковидной, нитевидной и сперматозоидной формы. Вирусная частица называется вирионом. Она состоит из нуклеиновой кислоты (ДНК или РНК) и белка глобулина; некоторые вирусы содержат также липиды и углеводы. Харак-
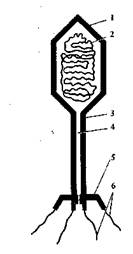
Рис. 18. Схема строения фага:
1 — головка; 2 — ДНК; 3 — отросток; 4 — стержень; 5 — базаль-ная пластинка с шипами; 6 — нити отростка
терной особенностью вирусов является их способность к образованию кристаллов, что долгое время служило причиной споров о живой или неживой природе вирусов. Впоследствии было доказано, что кристаллы — это нуклеиновая кислота и белок. Затем был установлен ряд свойств, подтвердивших представление о живой природе вирусов, — способность к самовоспроизведению (размножению), изменчивость, приспособляемость к условиям существования, а также способность вызывать инфекционные процессы. Развитие и размножение вирусов возможно только в клетках живого организма — хозяина, т. е. они являются паразитами человека, вызывая инфекционные заболевания (грипп, полиомиелит, корь, ветряная оспа и др.), а также животных и растений.
Для лечения некоторых заболеваний, вызванных вирусами гриппа, герпеса и аденовирусами, применяют ферментные препараты— нуклеазы, вызывающие разрушение нуклеиновых кислот, что лишает вирусы способности к самовоспроизведению, а следовательно, ликвидирует их инфекционность.
Вирусы могут быть паразитами не только человека, животных и растений, но и микроорганизмов —грибов, актиномице-тов, бактерий. Такие вирусы получили название фагов. Вирусы бактерий называются бактериофаги, актиномицетов — актино-фаги, грибов — микофаги.
В 1898 г. русский ученый Н. Ф. Гамалея при изучении сибирской язвы крупного рогатого скота впервые наблюдал, что спорообразующие палочки — возбудители болезни — растворяются под влиянием какого-то агента. В 1915 г. английским микробиологом Ф. Туортом и в 1917 г. канадским микробиологом Ф. Д'Эррелем была установлена природа этого явления. Оно получило название бактериофагии, а возбудитель — бактериофага («пожиратель бактерий»).
Размеры фагов колеблются от 40 до 140 нм. Бактериофаги имеют вид многогранной головки со стержнем, покрытой снаружи белковой оболочкой (рис. 18). Внутри стержня имеется канал. Головка фага заполнена молекулой ДНК. У основания стержня имеется базальная пластинка с шипами и нитями.
Воздействие фага на бактериальную клетку происходит в несколько стадий (рис. 19): адсорбция фага на бактериальной клетке с помощью базальной пластинки с зубцами и нитями, проникновение ДНК из головки фага по каналу в бактериальную клетку, в которой затем под влиянием фаговой ДНК
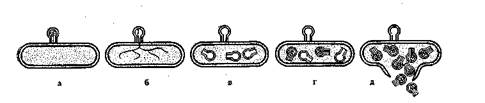
Рис* 19. Схема развития фага в бактериальной клетке:
а — адсорбция; б — переход ДНК в клетку; в — перестройка обмена веществ в клетке;
г — образование новых частиц бактериофага; д — растворение клеточной стенки
происходит полная перестройка обмена веществ, синтезируется уже не бактериальная ДНК, а фаговая, что приводит к образованию в бактериальной клетке новых частиц фага растворение клеточной стенки бактерии, ее гибель.
Бактериофаги наносят большой вред в молочной промышленности (производстве сыров, творога, сметаны) и в производстве маргарина. Они поражают в основном молочнокислые стрептококки заквасок для получения этих продуктов. Под влиянием бактериофага клетки стрептококков лизируются (растворяются) и погибают. В антибиотической промышленности актинофаги лизируют производственную культуру актиномицетов — продуцентов антибиотиков.
В медицине бактериофаги применяются для лечения некоторых заболеваний, например дизентерии.
Глава 2.
Дата добавления: 2015-11-20; просмотров: 28463;