Стебель - это осевой, преимущественно надземный орган растения, обладающий неограниченным ростом и выполняющий опорную, проводящую и запасающую функции.
Лекция. Первичное строение стебля.
1. Общая морфологическая характеристика стебля.
Стебель - это осевой, преимущественно надземный орган растения, обладающий неограниченным ростом и выполняющий опорную, проводящую и запасающую функции.
1) Стебель – осевой орган, обладающий радиальной симметрией (полисимметрией), то есть, через центр стебля можно провести много плоскостей симметрии.
2) Как правило, стебель – это надземный орган, то есть находится в воздушной среде. Бывают видоизменённые подземные стебли (у корневищ, клубней, луковиц).
3) Стебель несёт на себе листья, на нём располагаются почки, благодаря чему стебель ветвится.
4) Для стебля характерен практически неограниченный верхушечный ростза счёт апикальных меристем. Кроме того, стебель может расти и в ширину за счёт боковых меристем.
5) Стебель – это орган, связывающий корневую и листовую системы растения, поэтому его основные функции– это:
1. Проводящая
2. Механическая (опорная).
Именно благодаря проводящим тканям осуществляется нормальное функционирование всех органов растения.
Благодаря механическим тканям стебель выносит к свету листья, цветки и плоды.
Для стебля могут быть характерны и другие функции:
3. Запасающая – у деревьев в древесине развита запасающая паренхима. У некоторых степных и пустынных растений развивается водозапасающая ткань (кактусы, некоторые молочаи, баобаб, бутылочные деревья).
4. Для зелёных молодых стеблей характерна ассимиляционная функция.
Т.о, стебель – осевой орган растения (осевая часть побега), состоящий из узлов и междоузлий к которым прикрепляются листья, растущий за счёт интеркалярных, боковых и апикальных меристем.
6) По форместебель чаще всего бывает округлым (цилиндрическим), так как это наиболее экономично и обеспечивает максимальную структурную прочность.
7) Геотропизм стебля, т.е. чувствительность к силе земного притяжения, отрицательный, так как он растёт против силы земного притяжения. Это свойство является важной приспособительной функцией стебля.
 а) Обычно отрицательно геотропичен главный стебель, а боковые отходят под углом от него, что является важной адаптацией. Благодаря этому листья не затеняют друг друга.
а) Обычно отрицательно геотропичен главный стебель, а боковые отходят под углом от него, что является важной адаптацией. Благодаря этому листья не затеняют друг друга.
 б) Иногда отрицательным георопизмом могут обладать боковые стебли. Тогда формируется пирамидальная крона (кипарис, пирамидальные тополя).
б) Иногда отрицательным георопизмом могут обладать боковые стебли. Тогда формируется пирамидальная крона (кипарис, пирамидальные тополя).
 в) У некоторых растений боковые стебли обладают положительным геоторопизмом. Тогда формируется плакучая
в) У некоторых растений боковые стебли обладают положительным геоторопизмом. Тогда формируется плакучая
крона (берёза, некоторые ивы).
2. Анатомия стебля.
Чтобы понять, почему растёт стебель, рассмотрим строение его апекса.
Апекс (лат. apex – вершина) - это верхушка побега (стебля) или кончика корня, где расположены апикальные (верхушечные) меристемы.
Главная, наиболее продуктивная часть апекса – этоконус нарастания. У стебля конус нарастания представляет собой гладкую, наиболее удалённую от центра (дистальную (distans – max далеко)) часть апекса, до первых листовых бугорков. Здесь сосредоточены наиболее молодые меристемы (промеристемы) за счёт деления клеток которых происходит акропетальное, т.е. снизу вверх нарастание стебля.
У разных растений конус нарастания имеет разное строение и механизм нарастания, что привело к появлению различных теорий строения апекса.
Разберём особенности строения и жизнедеятельности конуса нарастания в разных систематических группах растений.
1.  Наиболее просто устроен конус нарастания у многоклеточных водорослей. Его строение было изучено Негели в 40-е годы 19 века. Водоросли нарастают за счёт деления одной единственной инициальной клетки, которая постоянно делится поперечными перегородками.
Наиболее просто устроен конус нарастания у многоклеточных водорослей. Его строение было изучено Негели в 40-е годы 19 века. Водоросли нарастают за счёт деления одной единственной инициальной клетки, которая постоянно делится поперечными перегородками.
В результате эволюции у высших споровых растений происходит усложнение работы инициальной клетки.
2.  У высших споровых растений в апексе имеется тоже только одна инициальная клетка, но деления её более сложные.
У высших споровых растений в апексе имеется тоже только одна инициальная клетка, но деления её более сложные.
У плауновинициальная клетка двухграннаяи еёделение идёт параллельнодвум граням.
Затем, в эволюции, деления инициальной клетки усложняются и возникает трёхгранная клетка (у хвощей и папоротников).
 Её деления более сложные. Она откладывает свои производные в трёх плоскостях параллельно трём граням.
Её деления более сложные. Она откладывает свои производные в трёх плоскостях параллельно трём граням.
Дальнейшая эволюция приводит к появлению в апексе не одной а многих инициальных клеток, которые у наиболее высокоорганизованных семенных растений образуют слои (или зоны).
3. Апексы голосеменных и цветковых растений имеют сложное строение, они состоят из различных по форме и типу делений клеток, формирующих разные ткани.
1) Впервые апексы семенных растений были изучены Ганштейном (60-е годы 19 века). По его схеме, точка роста (конус нарастания) семенных растений состоит из 3 слоёв клеток, принимающих участие в формировании разных постоянных тканей. (Причём, у покрытосеменных растений по сравнению с голосеменными, слоистость и первоначальная дифференцировка апикальных меристем выражена более чётко.) Его теория получила название – теория гистогенов.(Гистоген – это специализированная меристема, судьба которой предопределена, т.е. из неё формируются строго определённые ткани.)
Т.о., согласно Ганштейну, в конусе нарастания семенных растений выделяются следующие слои (зоны) инициальных клеток:
1 слой – дерматоген - наружный слой из одного ряда клеток (sin.= протодерма). Клетки делятся только поперечными перегородками перпендикулярно поверхности и не дают производных внутрь.. Впоследствии из дерматогена образуется эпидерма.
2 слой – периблема – 1-4 слоя клеток под дерматогеном. Она как колпачок покрывает внутренние слои. Её клетки делятся, в основном, перпендикулярно поверхности, реже тангентально (то есть иногда дают производные внутрь). Впоследствии из периблемы формируется первичная кора стебля.
3 слой – плерома – Это вся основная, осевая масса клеток конуса нарастания. Её клетки делятся во всех направлениях, формируя главную часть стебля – центральный цилиндр. На некоторой глубине, ниже конуса нарастания клетки плеромы дифференцируются в перицикл, прокамбий, формирующий проводящие пучки и сердцевину.
При заложении нового листа или ветви, клетки периблемы активно делятся и образуют бугорок, который заставляет вытягиваться и слой дерматогена (который играет пассивную роль). Клетки периблемы, лежащие под ним, затем дифференцируются на периблему и плерому.
2) Дальнейшие исследования, проведённые нем. ботаниками Дж. Будером и А. Шмидтом (1924 – 1928 г.) показали, что не всегда в конусе нарастания чётко видны все три зоны (например, граница между дерматогеном и периблемой часто совершенно незаметна). Поэтому была предложена новая теория роста стебля, получившая название теория туники-корпуса.
В конусе нарастания (точке роста) стебля имеется всего 2 комплекса клеток: туника и корпус.
1 комплекс – туника (греч. туника – одежда) – представлена 2-4 наружными слоями клеток со своими инициалями. Имеет вид колпачка, покрывающего внутренние слои, и обеспечивает поверхностный рост апекса, не давая внутрь своих производных. Из наружного слоя туники затем дифференцируется эпидерма, из нижних слоёв - первичная кора.
2 комплекс – корпус (лат. corpus – тело) – находится под туникой, имеет свои инициали. Клетки корпуса делятся во всех направлениях, обеспечивая объёмный рост апекса, однако, деления эти достаточно редки. Из него формируется, в основном, сердцевина стебля.
Но эта теория имела один существенный недостаток:она недостаточно чётко определяла из какого слоя, туники или корпуса образуются главные элементы стебля: проводящие и механические.
3) Поэтому американским ботаником Э.Форстером была предложена 3 теория цитогистологической зональности,объединившая и дополнившая две предыдущие теории. Её суть заключается в том, что в конусе нарастания есть туника и корпус. В нижней части конуса нарастания, там, где должен возникнуть очередной листовой зачаток (листовой примордий), деления клеток на границе этих двух зон резко усиливаются, внутренняя граница туники и внешняя граница корпуса сливаются и образуется зона периферической меристемы (или область инициального кольца). Здесь определяется порядок заложения листовых зачатков, почек и закладывается прокамбий (который затем формирует первичную флоэму и ксилему стебля). В центре стебля, внутрь от зоны инициального кольца, обособляется стержневая меристема, клетки которой делятся очень редко, формируя сердцевину стебля, состоящую из паренхимы. Самый наружный слой туники становится протодермой (зачаточной эпидермой будущего стебля). Внутренние слои туники формируют первичную кору стебля.
Прокамбий закладывается в зоне периферической меристемы на границе листового бугорка и оси побега в виде тяжей узких и длинных (прозенхимных) продольно делящихся клеток. Он распространяется и в растущий лист и вдоль оси побега, давая начало единой проводящей системе листа и стебля.
Протодерма и прокамбий являются гистогенами, то есть специализированными меристемами образующими строго определённые типы постоянных тканей – эпидерму, флоэму и ксилему соответственно.
Т.о., формируется стебель первичного строения, т. е. стебель, ткани которого сформировались благодаря деятельности первичных меристем.
Возникновение первичных проводящих тканей из прокамбия.
 В основании молодых листовых зачатков, в зоне инициального кольца закладывается прокамбий. Он может закладываться либо в виде сплошного кольца, либо в виде отдельных продольных тяжей. Ниже зоны инициального кольца прокамбий уже хорошо заметен, так как состоит из прозенхимных, удлинённых клеток. В ниже расположенных участках стебля, клетки прокамбия дифференцируются в элементы первичной флоэмы и ксилемы.
В основании молодых листовых зачатков, в зоне инициального кольца закладывается прокамбий. Он может закладываться либо в виде сплошного кольца, либо в виде отдельных продольных тяжей. Ниже зоны инициального кольца прокамбий уже хорошо заметен, так как состоит из прозенхимных, удлинённых клеток. В ниже расположенных участках стебля, клетки прокамбия дифференцируются в элементы первичной флоэмы и ксилемы.
Флоэма закладывается раньше ксилемы в наружных участках прокамбия, т.е. экзархно (греч. «экзо» - снаружи и «архе» - начало) и развивается центростремительно (т.е. по направлению к центру стебля). Ксилема закладывается из внутренних частей прокамбия , т.е. эндархно, и развивается центробежно (т.е. от центра стебля). Т.о., флоэма и ксилема развиваются в противоположных направлениях навстречу друг другу.
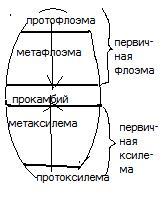 По времени образования и структурным особенностям первичная флоэма бывает двух видов: 1) протофлоэма
По времени образования и структурным особенностям первичная флоэма бывает двух видов: 1) протофлоэма
2) метафлоэма.
Протофлоэма – самая наружная, ранняя часть флоэмы. Состоит из удлинённых, тонкостенных, слабо дифференцированных живых клеток. Ближе к центру стебля, внутрь от протофлоэмы дифференцируется метафлоэма, содержащая хорошо развитые ситовидные трубки с клетками-спутницами, лубяные волокна и паренхиму. Она формируется позднее, после протофлоэмы.
Точно также, первичная ксилема представлена: 1) протоксилемой
2) метаксилемой.
Протоксилема развивается из самых внутренних участков прокамбия на границе с сердцевиной и состоит из спиральных и кольчатых сосудов и трахеид. Между проводящими располагаются паренхимные элементы. В метаксилеме, развивающейся позднее и ближе к поверхности стебля, клетки хорошо дифференцированы и проводящие элементы представлены лестничными, сетчатыми и точечно-поровыми сосудами или трахеидами.
Прокамбий или полностью расходуется или остатки прокамбия превращаются в камбий.
Дата добавления: 2015-08-14; просмотров: 1877;