Эритроциты
Эритроциты или красные кровяные тельца представляют собой безъядерные клетки двояковогнутой формы. Диаметр эритроцитов - 7-8 мкм, толщина - 2,5 мкм. В I мм3 крови находится 4-5 млн. эритроцитов. Общая поверхность эритроцитов взрослого человека составляет 3800 м2. Продолжительность жизни эритроцитов - 100-130 дней. Ежесекундно в красном костном мозге их образуется около 5 млн. Разрушаются они в селезенке и в печени. Их основная функция - перенос кислорода из легких в ткани и двуокиси углерода из тканей в легкие. Поступление веществ в эритроциты осуществляется либо вследствие диффузии, либо через поры, либо через особые участки мембраны.
Переносу кислорода способствует двояковогнутая форма эритроцитов: их поверхность при такой форме максимальна, что облегчает диффузию кислорода внутрь клетки. При такой форме диффузионная поверхность увеличивается, а диффузионное расстояние уменьшается, т.к. клетка становится более плоской. Благодаря подобной конфигурации клетка легко деформируется при прохождении через узкий изогнутый просвет капилляров, чему способствует эластичность мембраны. По мере старения клеток способность к изменению формы уменьшается. В связи с тем, что одна из главных функций эритроцитов - перенос кислорода, окислительные процессы в эритроцитах снижены, т.к. они перешли на бескислородное окисление - гликолиз. Отсутствие ядра у эритроцитов способствовало снижению уровня обмена веществ и улучшило возможность переноса кислорода.
5.1.1. Эритропоэз.
Эритроциты образуются в печени, селезенке у плода и красном костном мозге у взрослых. Родоначальные стволовые клетки являются предшественниками всех клеток крови. Из них в процессе дифференцировки образуются эритроциты, гранулоциты, тромбоциты или лимфоциты. Пройдя несколько этапов развития, из этих стволовых клеток образуются эритробласты, далее ретикулоциты. Последние представляют юные безъядерные эритроциты, которые выходят из костного мозга. В дальнейшем эти клетки созревают, и образуются эритроциты. За одну минуту образуется 160*106. При недостатке витамина В12 возникает мегалобластическая анемия, которая характеризуется наличием в крови прежде всего патологически увеличенных эритроцитов – мегалоцитов – и их незрелых предшественников – мегалобластов.
Физиологические механизмы регуляции эритропоэза состоят в следующем. Стимулом для эритропоэза может быть снижение парциального давления 02 или кровопотеря. Это ведет к образованию в почках эритропоэтина, который, в свою очередь, ускоряет размножение предшественников эритроцитов в костном мозге. Это ведет к увеличению числа эритроцитов и концентрации гемоглобина в крови в связи со стимуляцией костного мозга, где они образуются. Это имеет большое приспособительное значение, т.к. обеспечивает потребность человека в кислороде в условиях высокогорья. Количество эритроцитов в крови регулируется нервной системой и гуморальным путем. Действие эритропоэтина усиливается рядом гормонов - тироксином, гормоном роста и др.
Снижение способности крови переносить кислород называется анемией. Причинами анемии может быть уменьшение числа эритроцитов, количество гемоглобина. Анемия может быть связана с недостаточностью витамина В12 и железа в пищевых продуктах, нарушением всасывания в кишечнике, кровопотерями.
5.1.2. Транспорт кислорода
Кислород переносят молекулы гемоглобина, которые находятся в эритроцитах. В состав гемоглобина входит белок - глобин и четыре молекулы гема. Гемоглобин придает крови красный цвет. Каждый гем содержит атом двухвалентного железа, непрочно связывающий одну молекулу кислорода. Большая часть кислорода в крови находится в химически связанном состоянии.
Нв + 402= Нв(402). 1 моль гемоглобина может связать, таким образом, четыре молекулы кислорода. В ходе экспериментальных исследований было выяснено, что один грамм гемоглобина удерживает 1,34 – 1,36 мл 02. Зная содержание гемоглобина в одном литре крови, а оно равно 150 г, можно вычислить кислородную емкость крови. Она будет приблизительно равна
1.34 мл 02*150г/л Нв = 200 мл 02 на один литр крови. Это тот объем кислорода, который может теоретически раствориться в 1 л крови. В естественных условиях кислородная емкость крови несколько меньше. Было выяснено, что при прохождении крови через капилляры используется только 25% от общей кислородной емкости крови. Степень извлечения кислорода различными органами широко варьирует.
Образование оксигемоглобина, т.е. соединение гемоглобина с кислородом, происходит при высоком парциальном давлении кислорода в легких. При низком парциальном давлении (например, в капиллярах тканей) связь между гемоглобином и кислородом становится непрочной, кислород освобождается и диффундирует в окружающие ткани. Количество кислорода, которое может связаться с гемоглобином, зависит от уровня парциального давления. Парциальное давление – это давление газа в смеси газов, которое приходится на долю данного газа. Например, атмосферное давление на уровне моря составляет 760 мм.рт.ст. Кислорода в атмосфере воздуха содержится около 21%. Следовательно, его парциальное давление на уровне моря равно
760*21% = 152 мм. рт. ст
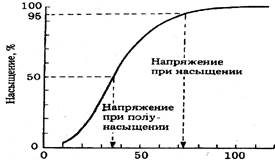 Гемоглобин можно считать полностью насыщенным кислородом при таком напряжении кислорода, при котором насыщается 95% гемоглобина. В легочных капиллярах парциальное давление меньше, чем в воздухе, но более 100 мм рт.ст., что достаточно для полного насыщения гемоглобина. Это позволяет организму не испытывать недостаток в кислороде при колебаниях парциального давления в легких в пределах от 70-100 мм рт. ст. Это фактор надежности. При более высокой концентрации кислорода существенного изменения насыщения уже не происходит. На графике видно, что при парциальном давлении, равном 73 мм рт.ст., происходит полное насыщение гемоглобина. При парциальном давлении кислорода 30 мм.рт.ст. только 50% гемоглобина находится в форме оксигемоглобина, а при нулевом парциальном давлении кислорода молекулы гемоглобина вообще не связывают кислород. В области крутого наклона кривой уже при небольшом снижении парциального давления кислорода процент насыщения гемоглобина значительно уменьшается. Увеличение способности связывания гемоглобина вызвано тем, что при взаимодействии кислорода с атомом двухвалентного железа одного из гемов структура его изменяется. Это, в свою очередь, изменяет конфигурацию всей молекулы гемоглобина. По мере присоединения кислорода ко второй и третьей группам гема происходит дальнейшее изменение в строении гемоглобина, в результате чего каждая молекула кислорода присоединяется легче, чем предыдущая, так что последний гем связывает кислород в сотни раз легче, чем первый. Также происходит и обратный процесс - диссоциация оксигемоглобина до гемоглобина. В области низкого парциального давления в активно работающих тканях первая молекула оксигемоглобина отщепляется очень легко, а далее этот процесс замедляется. При повышении парциального давления углекислого газа кривая диссоциации оксигемоглобина сдвигается вправо.
Гемоглобин можно считать полностью насыщенным кислородом при таком напряжении кислорода, при котором насыщается 95% гемоглобина. В легочных капиллярах парциальное давление меньше, чем в воздухе, но более 100 мм рт.ст., что достаточно для полного насыщения гемоглобина. Это позволяет организму не испытывать недостаток в кислороде при колебаниях парциального давления в легких в пределах от 70-100 мм рт. ст. Это фактор надежности. При более высокой концентрации кислорода существенного изменения насыщения уже не происходит. На графике видно, что при парциальном давлении, равном 73 мм рт.ст., происходит полное насыщение гемоглобина. При парциальном давлении кислорода 30 мм.рт.ст. только 50% гемоглобина находится в форме оксигемоглобина, а при нулевом парциальном давлении кислорода молекулы гемоглобина вообще не связывают кислород. В области крутого наклона кривой уже при небольшом снижении парциального давления кислорода процент насыщения гемоглобина значительно уменьшается. Увеличение способности связывания гемоглобина вызвано тем, что при взаимодействии кислорода с атомом двухвалентного железа одного из гемов структура его изменяется. Это, в свою очередь, изменяет конфигурацию всей молекулы гемоглобина. По мере присоединения кислорода ко второй и третьей группам гема происходит дальнейшее изменение в строении гемоглобина, в результате чего каждая молекула кислорода присоединяется легче, чем предыдущая, так что последний гем связывает кислород в сотни раз легче, чем первый. Также происходит и обратный процесс - диссоциация оксигемоглобина до гемоглобина. В области низкого парциального давления в активно работающих тканях первая молекула оксигемоглобина отщепляется очень легко, а далее этот процесс замедляется. При повышении парциального давления углекислого газа кривая диссоциации оксигемоглобина сдвигается вправо.
Напряжение 02 , мм рт. ст
Схема поясняет, что при напряжении 02 73 мм.рт.ст. 95% гемоглобина насыщаются кислородом. Напряжение полунасыщения - напряжение, при котором насыщено 02 50% пигмента.
.
Изменение способности связывать гемоглобином кислород играет большую роль в организме. В капиллярах работающих тканей количество углекислого газа в крови увеличивается, и поэтому уменьшается способность гемоглобина связывать кислород, что облегчает отдачу кислорода тканям. В альвеолах легких, где часть углекислого газа переходит в альвеолярный воздух, способность гемоглобина связывать
кислород возрастает, т.е. увеличивается образование оксигемоглобина. Резко снижается способность связывать гемоглобином кислород в крови мышечных капилляров во время интенсивной мышечной работы, когда в кровь поступает большое количество молочной кислоты. Это означает, что при увеличении водородных ионов в крови сродство гемоглобина кислороду уменьшается. Поэтому при увеличении углекислого газа при одном и том же парциальном давлении кислорода уменьшается образование оксигемоглобина.
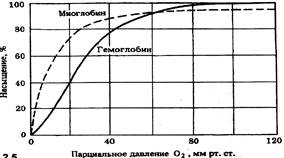 Миоглобин находится в мышцах животных, особенно в скелетных мышцах млекопитающих. Процент насыщения миоглобина кислородом происходит при более низких цифрах парциального давления кислорода. На графике видно, что при снижении парциального давления кислорода процент насыщения оксигемоглобина кислородом в сравнении с миоглобином меньше. Миоглобин остается насыщенным 02 на 80% до тех пор, пока парциальное давление кислорода не падает ниже 20 мм.рт.ст. (гемоглобин – на 40% !) Это означает, что миоглобин удерживает кислород в покоящемся мышечном волокне, но отдает его, когда в результате интенсивной мышечной работы используется весь кислород, доставленный гемоглобином. Т.е. миоглобин является резервуаром кислорода в мышцах.
Миоглобин находится в мышцах животных, особенно в скелетных мышцах млекопитающих. Процент насыщения миоглобина кислородом происходит при более низких цифрах парциального давления кислорода. На графике видно, что при снижении парциального давления кислорода процент насыщения оксигемоглобина кислородом в сравнении с миоглобином меньше. Миоглобин остается насыщенным 02 на 80% до тех пор, пока парциальное давление кислорода не падает ниже 20 мм.рт.ст. (гемоглобин – на 40% !) Это означает, что миоглобин удерживает кислород в покоящемся мышечном волокне, но отдает его, когда в результате интенсивной мышечной работы используется весь кислород, доставленный гемоглобином. Т.е. миоглобин является резервуаром кислорода в мышцах.
5.1.3. Транспорт углекислого газа кровью
В организме млекопитающих, и в том числе человека, существуют три способа переноса СО2:
А) 5% переносится в физически растворенном состоянии. Количество физически растворенного С02 в 9 раз больше, чем кислорода. Это связано с тем, что коэффициент растворимости для С02 в 20 раз больше, чем для О2. Несмотря на то, что количество С02 и 02 в состоянии физической растворимости невелико, эти газы должны пройти этап физической растворимости, чтобы перейти в химически связанное состояние.
Б) Примерно 25-30% углекислого газа, поглощаемого в капиллярах большого круга кровообращения, вступает в соединение с гемоглобином, образуя карбогемоглобин.
В) Большая часть - до 80% - переносится в форме карбонатов. Последний процесс происходит следующим образом. Образующийся С02 в тканях диффундирует в кровь, так как концентрация его в тканях больше, чем в крови. Далее он диффундирует в эритроциты, где под влиянием фермента карбоангидразы превращается в угольную кислоту (СО2 + Н20 = Н2СО3). Угольная кислота диссоциируется на Н+ и НСО3–. Часть НСО3– диффундирует в плазму, где соединяется с Na+ и в форме бикарбоната натрия NaНСО3 транспортируется плазмой крови. Другая часть НСО3– в эритроцитах соединяется с ионом К+ и транспортируется эритроцитами в форме бикарбоната калия КНСО3.
Дата добавления: 2015-07-14; просмотров: 4057;