Влияние условий на интенсивность транспирации
Как степень открытия устьиц влияет на интенсивность транспирации, так же внешние условия влияют на транспирацию , как на физический процесс.
Прежде всего на транспирацию влияет степень насыщенности атмосферы водой. Чем меньше относительная влажность воздуха, тем ниже его водный потенциал и тем быстрее идет транспирация. С увеличением влажности воздуха транспирация уменьшается; и при большой влажности воздуха происходит только гуттация
Транспирация зависит от температуры. С её повышением значительно увеличивается количество воды, насыщающей воздушное пространство, что увеличивает дефицит влаги в воздухе, а следовательно увеличивается и транспирация.
Свет – главный фактор, регулирующий транспирацию, который на испарение воды с водной поверхности практически не влияет. Влияние света на транспирацию связано прежде всего с тем, что клетки листа, содержание хлорофилл, поглощают не только инфракрасные лучи, но и видимы свет, необходимый для фотосинтеза. На фотосинтез лист расходует только 1-2% поглощенного света, а остальная энергия (98%) превращается в тепло, которая используется на испарение. Свет так же косвенно влияет на транспирацию, вызывая открытие устьиц.
В естественных условиях на интенсивность транспирации влияет ветер, но не так сильно как на испарение со свободной водной поверхности. В основном его влияние проявляется на третьем этапе устьичной транспирации, так как действие на внутренние ткани, из которых идет испарение, ограничено. Увеличение интенсивности транспирации происходит до 0,4 – 0,8 м/с, дальнейшее усиление ветра уже не влияет на транспирацию, так как в таком случае транспирация зависит от диффузии воды через устьица.
Транспирация зависит от влажности почвы. Здесь прямая зависимость. Чем меньше воды в почве, тем меньше её в клетках листа, а следовательно транспирация уменьшается., так как устьица прикрываются. Транспирация так же снижается, при недостатке недостаток кислорода в почве, что снижает дыхание корней, поглощение ими минеральных веществ, а следовательно и воды.
Интенсивность транспирации зависит от условий минерального питания, При недостатке азота, фосфора, или калия усиливает транспирацию. Транспирация сокращается
если растение получает полное минеральное удобрение. Это объясняется тем, что поглощенные ионы и синтезированные вещества связывают воду, а испаряется только свободная вода.
Транспирация зависит и от внутренних условий.
Прежде всего интенсивность транспирации зависит от содержания воды в листьях. Всякое уменьшение воды в листьях уменьшает транспирацию.
Интенсивность транспирации зависит от количества свободной воды, водоудерживающей силы клеток. Чем меньше свободной воды, и меньше транспирация.
Величина листовой поверхности так же является условием влияющим потери воды растением. Чем она больше, тем и больше теряется воды. Однако здесь могут быть различные приспособления растений, выработанных в процессе эволюции, снижающих испарение: опушенность листовой пластинки, погружение устьиц в мезофилл, толщина кутикулы.
Интенсивность транспирации зависит от соотношения поверхности корней и побегов. При увеличении доли корней транспирация увеличивается.
Возрастные изменения как листа, так и растения существенно влияют на интенсивность транспирации. Она может меняться у разных растений в зависимости от фаз и этапов онтогенезе.
ПЕРЕДВИЖЕНИЕ ВОДЫ ПО РАСТЕНИЮ.
| Рис. Схема расположения концевых двигателей водного тока – паренхима листа и корня – и соединяющих их сосудов. |
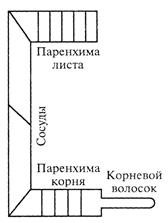 .Высшие растения расчленены на органы, выполняющие различные функции, Листья синтезируют органические вещества и нуждаются в поступлении притока воды и содержащихся в ней минеральных веществах. Корни поглощают воду и минеральные вещества, а в свою очередь нуждаются в постоянном притоке органических веществ, необходимых для дыхания и процессов жизнедеятельности, которые в них не образуются, корни в отношении органических веществ гетеротрофные. Это требует постоянного передвижения воды по растению. У растений сформировалось два транспортных потока – восходящий ток, по которому вода с растворенными в ней минеральными и органическими веществами , образуемых в корне( АТФ, аминокислоты, фитогормоны) движется от корня во все части наземных органов. Ложем для него служат полые сосуды ксилемы: трахеи и трахеиды, мертвые, не содержащие протоплазмы, которые являются основной частью апопласта. Апопласт – это совокупность свободного водного пространства клеточных стенок и межклетников. И второй ток – ток пластических веществ, который движется по живым клеткам флоэмы, ситовидным трубкам от мест синтеза органических веществ( листьев) к местам их потребления и в корень (вниз), к новообразованиям (молодые листья, генеративные органы, созревающие семена, точки роста др),т.е в отличии от водного восходящего тока, в разных направлениях. Поэтому называть нисходящим током передвижение веществ по флоэме не совсем верно. Его ложем служат живые клетки проводящей ткани флоэмы – ситовидные трубки, являющиеся частью системы – симпласта..
.Высшие растения расчленены на органы, выполняющие различные функции, Листья синтезируют органические вещества и нуждаются в поступлении притока воды и содержащихся в ней минеральных веществах. Корни поглощают воду и минеральные вещества, а в свою очередь нуждаются в постоянном притоке органических веществ, необходимых для дыхания и процессов жизнедеятельности, которые в них не образуются, корни в отношении органических веществ гетеротрофные. Это требует постоянного передвижения воды по растению. У растений сформировалось два транспортных потока – восходящий ток, по которому вода с растворенными в ней минеральными и органическими веществами , образуемых в корне( АТФ, аминокислоты, фитогормоны) движется от корня во все части наземных органов. Ложем для него служат полые сосуды ксилемы: трахеи и трахеиды, мертвые, не содержащие протоплазмы, которые являются основной частью апопласта. Апопласт – это совокупность свободного водного пространства клеточных стенок и межклетников. И второй ток – ток пластических веществ, который движется по живым клеткам флоэмы, ситовидным трубкам от мест синтеза органических веществ( листьев) к местам их потребления и в корень (вниз), к новообразованиям (молодые листья, генеративные органы, созревающие семена, точки роста др),т.е в отличии от водного восходящего тока, в разных направлениях. Поэтому называть нисходящим током передвижение веществ по флоэме не совсем верно. Его ложем служат живые клетки проводящей ткани флоэмы – ситовидные трубки, являющиеся частью системы – симпласта..
Передвижение воды по растению в восходящем токе можно разделить на 3 различные по структуре, физиологическим свойствам и роли в передвижении воды, участка:
1.Движение воды по живым клеткам корня до сосудов ксилемы (смотри рис. ),участок составляющий всего не более 3-х мл.
2. Движение по мёртвым сосудам ксилемы, достигающего от нескольких см до 150 м у древесных высокорослых растений.
3. По живым клеткам паренхимы мезофилла листа от проводящего пучка жилок до устьичной полости, в зависимости от толщины листа от 1 мм до не более 5 мм, или даже в несколько клеток( рис. ).
Как видим величина этих участков практически несоизмерима. В листьях и корне вода передвигается по апопласту и симпласту В стебле только по апопласту – мертвым сосудам ксилемы. Однако в стебле вода может передвигается от сосудов ксилемы в радиальном направлении по живым клеткам сердцевинных лучей, т.е по симпласту к клеткам камбия и живым паренхимным клеткам коры. Надо отметить , что вода по апопласту передвигается в наибольших объемах и достаточно быстрое ,не испытывая большого сопротивления, тогда как в симпласте это передвижение замедленное, связанно с препятствием преодоления сопротивления вязкой цитоплазмы.
Какие же силы способны преодолеть такие расстояния и обеспечить непрерывный ток воды между всеми органными растения? В основе механизма передвижения воды разница градиента водного потенциала между листьями и корнем. В корне, как уже отмечалось выше, движение воды связано с увеличением водного потенциала от корневого волоска до сосудов ксилемы, т. е. он ставится все боле отрицателен в результате чего вода поступает в сосуды ксилемы под давлением, получившего название корневого давления Оно не велико и составляет в среднем около 3 атм ( 0,3 кПа), способность поднять воду не выше 30 см.

 Передвигаясь пасока по сосудам попадает в лист через черешок. В листовой пластинке водопроводящие сосуды разветвляются, становясь все мельче и мельче, а следовательно вода встречает меньшее сопротивление и подводят воду к клеткам мезофилла. В мезофилле листа от жилок вода передвигается от клетки к клетке, в силу создаваемой разницы градиента водного потенциала, создаваемого испарением воды клетками, прилегающих к полостям устьиц, из которых воду уходит в атмосферу, создавая ненасыщеность клеточных стенок мезофилла, которое передается во внутрь клетки и их водный потенциал возрастает, т.е он становится более отрицательным, который передается к клетками прилегающим к сосудам. Этот отрицательный водный потенциал передается сосудам. из которых вода поступает в клетки мезофилла. Создается сильная отрицательная присасывающая сила листьев, создаваемая транспирацией.
Передвигаясь пасока по сосудам попадает в лист через черешок. В листовой пластинке водопроводящие сосуды разветвляются, становясь все мельче и мельче, а следовательно вода встречает меньшее сопротивление и подводят воду к клеткам мезофилла. В мезофилле листа от жилок вода передвигается от клетки к клетке, в силу создаваемой разницы градиента водного потенциала, создаваемого испарением воды клетками, прилегающих к полостям устьиц, из которых воду уходит в атмосферу, создавая ненасыщеность клеточных стенок мезофилла, которое передается во внутрь клетки и их водный потенциал возрастает, т.е он становится более отрицательным, который передается к клетками прилегающим к сосудам. Этот отрицательный водный потенциал передается сосудам. из которых вода поступает в клетки мезофилла. Создается сильная отрицательная присасывающая сила листьев, создаваемая транспирацией.
| Рис. Присасывающее действие испарения воды гипсом и транспирации |
Таким образом по сосудам вода движется благодаря присасывающей силы транспирации , создающей большую разницу градиента водного потенциала. Следовательно обнаруживается два мощных двигателя водного тока:
1) корневое давление и 2) присасывающая сила листьев.
Однако корневое давление. имеет величину 2-3 атм и его силы не достаточно , чтобы поднять воду выше 30 см. У деревьев же мы наблюдаем поднятие воды на сотни метров: эвкалипт, секвойя, сосна, лианы тропических лесов. Что же заставляет воду так легко и так высоко двигаться по сосудам. Оказалось , что в этом большую роль играют силы сцепления молекул воды в сосуда. В сосудах вода представляет собой неразрывную натянутую нить подвешенную к клеткам листа, и удерживаемую присасывающей силы транспирации. Что силы сцепления жидкости могут заставить её подниматься на высоту, превышающую барометрическую, давно показано опытом Аскеназы (1895), Он брал длинную стеклянную трубку с припаянной наверху воронку, залитую гипсом, и заполненную прокипяченной водой, лишенной воздуха, с водой трубки. Низ трубки он поместил в сосуд с ртутью, чтобы во всей системе не было пузырьков воздуха. С поверхности гипсовой воронки испаряемая вода втягивала воду из трубки, поднимая ртуть, которая поднималась несколько выше барометрической вследствие сцепления частиц воды между собой и ртутью ( рис. )
В таком состоянии вода в сосудах находится как бы в подвешенном состоянии к испаряющим клеткам листа виде тонких непрерывных нитей. Нижним Концом упираются в клетки корня, создающих корневое давление. Непрерывность и неразрывность водной нити создается силой сцепления молекул воды друг с другом и отсутствием пузырьков воздуха в них. Сцепление молекул воды друг с другом силой притяжения называется когезией.
С другой стороны молекулы воды водной нити удерживаются силой прилипания воды к стенкам сосуда, которая достигает200-300 атм (20-30 кПа).Связь молекул воды с гидрофильными стенками сосуда называется адгезия. Можно полагать , что микроскопический диаметр сосуда, способствуют тому, что силы сцепления преодолевают силу гравитационного веса водной нити.
Согласно теории сцепления вода в капиллярных трубках сосудов ксилемы поднимается вверх на присасывающую силу транспирации в результате действия сил сцепления молекул воды одной с другой (когезии) и действия сил прилипания ствола воды к гидрофильным стенкам сосудов( адгезии). Эти силы также препятствуют образованию пустот из воздуха, способных закупоривать сосуды (эмболии), приводящее к разрыву нитей воды и в других сосудах и прекращению передвижения воды. Сопротивлении воды разрыву составляет 300 атм (30 МПа), что обеспечивает подъем воды на 120-130 м,
Таким образом, механизм передвижения воды по растению можно рассматривать следующим образом: под действием корневого давления пасока или вода выталкивается в трахеи и сосуды ксилемы. Под действием транспирации создается присасывающая сила листьев, которая тянет водную нить, прочно удерживаемую силой молекулярного сцепления. В этой системе корень называют нижним концевым двигателем водного тока, а транспирацию листьев – верхним концевым двигателем. Скорость передвижения воды по сосудам у разных растений разная. У древесных растений лиственных пород (дуб) она достигает до 43,6 см/ час, у хвойных 12-14 0,5-1 см/ час.
Движущей силой ,как видим, восходящего сока в проводящих системах ксилемы является градиент водного потенциала через растение от почвы до атмосферы. Оно поддерживается:1) градиентом осмотического потенциала в клетках корня ( от почвы до сосудов ксилемы) в результате активного транспорта ионов в живых клетках и 2) транспирации, создающей отрицательный водный градиент в клетках листа. Поддержание первого градиента требует затраты метаболической энергии, второго использованием солнечной энергии.
ЗНАЧЕНЕ ВОСХОДЯЩЕГО ПОТОКА ВОДЫ ДЛЯ РАСТЕНИЙ 1) Восходящий ток от корневой системы до надземной части служит способом транспортирования и накопления в надземных органах минеральных веществ и химических соединений корня2) Транспирация связана с усвоением СО2.Чтобы получить СО2 растение должно отдавать воду, а уменьшение потери воды ( закрывание устьиц) уменьшает и приток Со2В сельском и лесном хозяйствах, для получения максимальных урожаев важно знать взаимоотношения между продукцией фотосинтеза и потери воды у растений с целью их регулирования.3) Восходящий поток необходимый для нормального водообеспечения всех клеток и под- держания тургора. При недостатке воды в клетках происходят различные нарушения. Поэтому для получения высоких урожаев в засушливых районах необходимо развивать орошаемое земледеиле.
ВЛИЯНИЕ ВОДНОГО ДЕФИЦИТА НА ФИЗИОЛОГИЧЕСКИЕ ПРОЦЕССЫ РАСТЕНИЙ.
При нарушении водного баланса при наступлении воздушной или почвенной засухи у растений появляется водный дефицит. Водный дефицит в растениях действует на такие процессы как: поглощение воды, транспирация, корневое давление, прорастание семян, фотосинтеза, дыхание, ферментативная активность, рост и развитие, взаимоотношение минеральных веществ. Изменяя обмен веществ, недостаток влаги влияет на продуктивность растений, вкус плодов, плотность древесины, длину и прочность волокна и.т.д. Содержание воды, необходимой для прорастания, различно для разных видов. Влияние водного дефицита на метаболические процессы зависит от её продолжительности. При затянувшемся завядании увеличивается скорость распада белков, нуклеиновых кислот. При этом содержание белка снижается в листьях и увеличивается в семенах .При водном дефиците (В.Д) содержание сахаров сначала уменьшается через снижение фотосинтеза, затем несколько возрастает в результате гидролиза полисахаридов листьев нижних ярусов и затем снова уменьшается ( все формы).В.Д снижает интенсивность фотосинтеза и образования АТФ, а также нарушает отток продуктов фотосинтеза листьев. При водном дефиците снижается дыхательный коэффициент, интенсивность дыхания резко падает, особенно у молодых листьев. В условиях водного дефицита. верхние листья увеличивают содержание осмотически активных веществ, оттягивают воду от нижних листьев и дольше сохраняют синтетические процессы в норме. В аридных зонах важно знать все физиологические особенности растений, и их изменения при В.Д., чтобы правильно определить сроки полива и их продолжительность.
ОСОБЕННОСТИ ВОДНОГО ОБМеНА у растений различных экологических групп.
1.Растения которые растут в воде, регулируют постоянство внутренней среды с помощью механизмов защиты от избыточного постоянного поступлениия воды, которую они поглощают всей поверхностью. 2.У наземных растений механизмы регуляции водного баланса направлены на защиту от значительной потери воды. Они разные у растений разных экологических групп. Под способностью приспосабливать водный обмен до колебаний водообеспечения различают 5 групп растений: 1.Пойкилогидрофиты (бактерии, сине-зеленые водоросли, зеленые водоросли порядка Protococcoales, грибы, лишайники, злаки сухих степей, пыльцевые зерна и семена покрытосеменных) приспособились переносить значительный недостаток воды без утраты жизнеспособности. При этом у них снижается обмен веществ. По характеру изменений всех показателей водного режима (осмотическое давление, интенсивность транспирации, содержание воды) на протяжении суток они относятся до гидролабильных растений.
2. Гомогидрофиты(наземные папоротникообразные, голосеменные, цветковые)имеют тонкие механизмы регуляции устьичной и кутикулярной транспирации, а также деятельности корневой системы. У всех клеток развита вакуолярная система и они не способны к обратимому высыханию. Показатели водного режима характеризуют гидростабильный тип. Делятся на 3 экологические группы.
3.Гигрофиты– тонколиственные папоротники, некоторые фиалки, калужница и др. – растения высокой влажности и /или затенения. Для них характерно : открытые устьица, гидатоды для выделения воды, плохо переносят какую-либо засуху.
4. Мезофиты –лиственные деревья, лесные и луговое травы, большинство культурных растений и т д. - занимают промежуточное положение между 5 и 7 группами.
5. Ксерофити – молочаи, алое, кактусы, полыни, ковыли и др.– распространены в местах с сухи жарким климатом, хорошо приспособлены к засухе.
Физиологические основы орошения земледелия
Рост и развитие растений очень сильно зависит от доступности воды.1/3 поверхности суши имеет недостаток влаги, половина из неё – крайне засушливы. Излишек влаги приходится лишь на 9% суши.
Когда годовые осадки превышают испарение, то говорят о гумидной зоне, в противоположном случае – о аридной. Земледелие в аридных зонах возможно только лишь при искусственно орошении.
ТЕОРИЯ ВОДНОГО РЕЖИМА И ОРОШЕНИЯ С/Х РАСТЕНИЙ.
Засухи и суховеи приводят к большим потерям в сельском хозяйстве. Сущность искусственного орошения заключается в установлении рационального поливного режима и системы питания растений в разных почвенно-климатических зонах
В этом контексте важным вопросом является верхняя и нижняя граница доступной влажности почвы ( ВГДВП, НГДВП)
ВГДВП(верхний градиент доступной воды почвы)называют полевую влагоёмкость. Когда влажность больше чем ВГ, то, несмотря на её доступность, она малоэффективна из-за недостатка кислорода. При НГДВП (нижний градиент доступной воды почвы) растения остро страдают от засухи, что сказывается на всей их жизнедеятельности. Влажность устойчивого завядания не может служить диагностическим показателем для определения сроков и норм полива, потому что снижение урожайности при почвенной засухе обусловлено не завяданием растений, а постоянным водным дефицитом в их тканях, которое наступает задолго до появления устойчивого завядания. Водный дефицит в тканях растений появляется при такой влажности почвы, при которой его водоудерживающая сила препятствует полному насыщению растений. Степень доступности почвенной воды в интервале от полной влагоёмкости до влажности устойчивого завядания для растений не одинакова. Поэтому важно определить то состояние влажности почвы, ниже которого нарушаются нормальные физиологические процессы. Такое состояние влажности почвы называют нижней границей оптимальной влажности (НГОВ)Величина НГО почвы зависит от фаз развития растений, особенностей почвенно-гидрологических и климатических условий, уровня агротехники и запланированной величины урожая, от наличия критических периодов с острой недостачей воды. Так для хлопчатника на типичных сероземах эти величины составляют до цветения – 75% полной влагоёмкости, в период цветения – 70%, а созревания –60% полной влагоёмкости. При этом существуют сортовые отличия.В производственных условиях сроки полива определяют:по влажности почвы ( на глубине 30-40 см.,по состоянию растений – уровню оводненности листьев, их осмотическому давлению, от степени открытости устьиц, способности выделять пасоку, по сосущей силе листьев. На данное время известны критические величины сосущей сили листьев для разных культур на разных фазах их развития. Они даны в справочниках. Своевременный полив приводит к повышению урожаю, Разработка физиологических показателей, которые дают возможность точно и экономно поливать растения в условиях засушливых зон – обязательные элементы высокой культуры поливного земледелия.
Дата добавления: 2015-08-11; просмотров: 5131;