ТРАНСПИРАЦИЯ
В основе расходования воды растительным организмом лежит физический процесс испарения – переход воды из жидкого состояния в парообразное, происходящее в результате соприкосновения органов растения с ненасыщенной водой атмосферой. Но у растений это не просто испарение воды, это сложный физиологический процесс регулируемый самим растением рядом анатомических и физиологических механизмов. Поэтому термин «транспирация» введен для того, чтобы подчеркнуть отличие этого физиологического процесса от чисто физического процесса. Транспирация это испарение воды с внутренних тканей листа через устьица. Таким образом, транспирация – это физиологический процесс испарения воды растениями. Главный орган транспирации – лист. Каковое же биологическое значение транспирации? Во-первых, транспирация обеспечивает непрерывность водного тока по растению его непрерывность, связывая растение в единое целое. Во-вторых. обеспечивает с водным током передвижение минеральных и частично органических веществ по растению , от корня по стеблю к листьям.
В третьих, регулирует температуру растения, защищает его от перегрева на прямых солнечных лучах. Без транспирации происходил бы быстрый перегрев растения, что вызывало бы нарушало биохимических и физиологических процессов. Транспирацию можно считать приспособительным процессом, возникшим в процессе эволюции растений после выхода на сушу. У водных растений, транспирации нет, и нет специальных органов транспирации – устьиц. Следовательно для растений она не обязательна, как, например, дыхание. Она наблюдается только у наземных и надводных растений.
Транспирация как физиологический процесс в свою очередь тесно связана с другими важнейшими процессами: фотосинтезом, дыханием, минеральным питанием. Так как через одни и те же органы – устьица идет газообмен и фотосинтеза,, и дыхания, а с минеральным питанием связано с транспортом веществ от корня с транспирационным током ко всем наземным органам. Как физиологический процесс транспирация имеет количественные и качественные единицы измерения, раскрывающие её напряженность и связь с фотосинтезом. Интенсивност транспирации, это количественный показатель, обычно выражают в граммах испарения воды за один час на единицу S или на один грамм сухой массы
( днем – 15-250 г/м2 за час; ночью – 1-20 г/м2 за час. Продуктивность транспирации, это качественный показатель, определяется количество граммов сухого вещества, образуемого при испарении 1000 граммов воды (≈ 1-8 г на 1000 г воды) Транспирационный коэффициент – количество граммов воды, которая затрачивается на образование одного грамма сухого вещества (≈ 120-150 г на 1 г сухого вещества Для оценки эффективности использования воды определяется коэффициент водопотребления – эвапотранспирация. Под эвапотранспирацией понимают суммарный расход воды за вегетационный период 1 га посева или насаждения. Сюда включается испарение как с почвенной поверхности (эвапорация) и транспирации всех растений, куда входят и сорные. Это можно использовать и при определении водопотреблении фитоценоза. Водопотреблением посева можно управлять, снижая расход воды путем уничтожения сорных растений, мульчированием почвы, своевременным рыхлением, улучшая продуктивность растений применение минеральных удобрений и других агротехнических приёмов Эта единица измерения водопотери используется при расчете выращивание растений при поливах..На синтез затрачивается всего 0,2% Н2О; остальная – на транспирацию
Лист как орган транспирации Основным транспирирующим органом растения является лист, общее строение листа известно из курса ботаники. Нам следует остановиться на его особенностях строения, связанных с транспирацией. Несмотря на то, что лист служит органом испарения воды ( транспирации) его анатомо-морфологичекое строение направлено на препятствие непродуктивной потери воды (рис ). Сверху и снизу лист покрыт эпидермисом, клетки которого плотно прижаты друг к другу, поверхность этих клеток покрыта кутикулой, непроницаемой для воды, к тому же она еще дополнительно покрывается восковым налетом. Кутин и воск – гидрофобны, что затрудняет испарение воды.
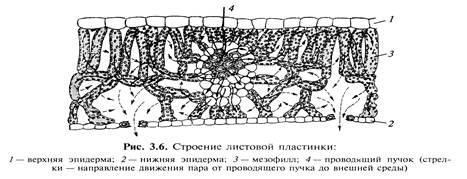
Несмотря на весьма небольшую толщину листа (100-200 мкм, строение мезофилла довольно сложное, которое определяет два важнейших процесса: фотосинтез и транспирацию. Эти процессы тесно взаимосвязаны. Их протекание определяется непосредственным контактом с атмосферой. Поэтому мезофилл имеет систему межклетников и проводящих пучков, первые увеличивают испаряющую поверхность и газообмен, вторые обеспечивают скорость поступления воды в листовую пластинку и отток пластических веществ, образуемых при фотосинтезе. Для контакта мезофилла с атмосферой в эпидермисе имеются мельчайшие отверстия устьица, через которые идет поглощение углекислого газа и испарение воды. Подсчеты показали, что площадь отверстий устьиц на поверхности листа составляет всего 1%, но самих устьиц на 1 мм2 приходится от 50 до 500 штук, т.е. весь лист в порах, но величина этих пор составляет всего – 3-12 мкм (нм). Обычно у наземных двудольных растений устьица расположены на нижней стороне листа, а у плавающих листьев на верхней . У растений лилейных и злаковых устьица расположены на обеих сторонах. Но оказалось , что если у листа двудольного растения замазать нижнюю сторону вазелином, то обнаруживается испарение воды с верхней стороны. Это испарение получило название кутикулярной транспирации. Таким образом, различают два основных типа транспирации: устьичную и кутикулярную транспирации. Некоторые ученые отмечают испарение воды через чечевички молодых побегов, и назвали его лентикулярной транспирацией. В результате транспираци чечевичек (лентикулярная транспирация) в зимнее время часто возникает водный дефицит и растения погибают через обезвоживание. По объему испарения воды, устьичная составляет до 90%, а кутикулярная – до 10%. Однако это соотношение не постоянно. В молодых растущих листьев, когда устьица еще полностью не сформировались, а кутикула тонкая – кутикулярная транспирация достигает 60 % и выше. По мере старения листа и развития кутикулы доля её в транспирации уменьшается. Более высокий процент кутикулярной транспирации наблюдается у теневыносливых растений, у которых кутикула слабо развита. Устьичная транспирация складывается из 3-х этапов:1) испарение воды из поверхности влажных клеток мезофилла; 2) диффузия паров воды через устьица и 3) диффузия водяных паров от поверхности листа (рис. ). Каждый этап имеет свои особенности и механизм регуляции транспирации.
Первый этап. Как известно, клеточная стенка представляет переплетение нитей микро- и макрофибрил, между которыми пространство заполнено водой, и вода находится как в капилляре. Вода испаряется из эти капилляров. При недостаточном снабжении листа водой, клеточные стенки подсыхают, мениск в капиллярах становится вогнутым, увеличивается поверхностное натяжение и удержание воды силами сцепления, испарение воды уменьшается. К тому же в подсохшей клеточной стенке вода уже испаряется не в межклетники, а во внутрь капилляра, в которых повышается давление паров, что так же снижает испарение воды. Мы видим регуляцию транспирации при открытых устьицах, Этот процесс получил название внеустьичная транспирация.
| Рис. Испарение и диффузия водяных паров в листе |
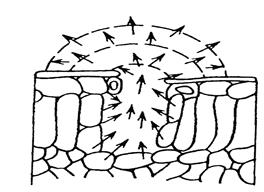 Второй этап начинается, когда вода отрывается от оболочек клеток мезофилла и переходит в межклетники и подустьичную полость в виде пара. Теперь пары будут устремляться к устьичной щели и проходить через неё. Под устьицем создается недостаток паров, что приводит автоматически к испарению воды с поверхности клеток. Иная картина наблюдается когда устьица закрыты, все пространство под ними быстро насыщается парами и переход воды из жидкого состояние в парообразное прекращается и потери воды листом нет. Вот поэтому степень открытия устьиц является основным механизмом регуляции интенсивности транспирации.
Второй этап начинается, когда вода отрывается от оболочек клеток мезофилла и переходит в межклетники и подустьичную полость в виде пара. Теперь пары будут устремляться к устьичной щели и проходить через неё. Под устьицем создается недостаток паров, что приводит автоматически к испарению воды с поверхности клеток. Иная картина наблюдается когда устьица закрыты, все пространство под ними быстро насыщается парами и переход воды из жидкого состояние в парообразное прекращается и потери воды листом нет. Вот поэтому степень открытия устьиц является основным механизмом регуляции интенсивности транспирации.
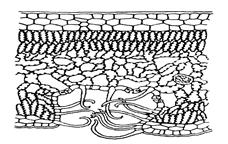 Третий этап устьичной трансирайии – это диффузия паров от поверхности листа. Этот этап регулируется в основном условиями внешней среды : температурой, влажностью воздуха, ветром, и приспособлениями, сокращающими транспирацию: опушение, погружению устьиц в мезофилл ( рис. ), скручивание листьев ( ковыль) и др.
Третий этап устьичной трансирайии – это диффузия паров от поверхности листа. Этот этап регулируется в основном условиями внешней среды : температурой, влажностью воздуха, ветром, и приспособлениями, сокращающими транспирацию: опушение, погружению устьиц в мезофилл ( рис. ), скручивание листьев ( ковыль) и др.
| Рис. Погружение устьиц в мезофилл и опушение |
Основное испарение воды идет через устьица. При полностью открытых устьицах транспирация идет интенсивно и почти приравнивается к испарению воды со свободной поверхности такой же площади. Но вспомним, что при полностью открытых устьицах их общая поверхность составляет всего 1% от площади листа, т.е казалось бы , что транспирация должна быть в 100 раз меньше, чем испарение со свободной поверхности воды. Но измерение транспирации и испарения с одинаковой водной поверхности показало, что транспирация всего меньше в 2 раза. Для сравнительной активности транспирации введено понятие относительной транспирации, которая показывает, что транспирация не только более интенсивна, но и саморегулируемая
.Относительная транспирация – это отношение транспирации к свободному испарению с водной поверхности той же площади и за то же времени. В среднем она составляет 0,2 -0,5, а у некоторых даже 0,9. Почему же так ?
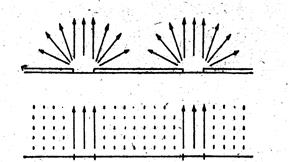
Объяснение этому дали в 1900 году английские физиологи Браун и Эскомби, которые установили, что испарение из ряда мелких отверстий идет быстрее , чем и одного крупного той же площади. Согласно Закону Стефана, связанного с так называемым краевым эффектом. Испарение с краёв идет быстрее, чем в центре. В центре происходит насыщение паров сильнее и связь с молекулами прочнее, а с краёв вода легче отрывается и скорее рассеивается, вокруг краев создаётся свободное диффузное поле и пары расходятся веером ( рис. ). Поэтому при испарении таких краевых молекул значительно больше у ряда мелких отверстий, расположенных друг от друга не ближе 10- кратного расстояния их диаметра, так как испарение пропорционально не площади, а диаметру отверстия
| Рис. Схема, объясняющая краевой эффект: маленькие отверстия (вверху), например устричная щель, имеют большее диффузионное поле, чем равная площадь открытой водной поверхности ( по Э. Либерту) |
 Устьица идеально отвечают этим требованиям: имеют малый диаметр и расположены на достаточном удалении друг от друга. При открытых устьицах испарение идет интенсивно, тогда как при их закрытии испарение снижается. В этом и заключается устьичная регуляция транспирации. При недостатке влаги устьица автоматически закрываются.
Устьица идеально отвечают этим требованиям: имеют малый диаметр и расположены на достаточном удалении друг от друга. При открытых устьицах испарение идет интенсивно, тогда как при их закрытии испарение снижается. В этом и заключается устьичная регуляция транспирации. При недостатке влаги устьица автоматически закрываются.
Основным фактором, обусловливающим движение устьиц, является содержание воды в листе. При достаточном её количестве, устьица открыты, при недостатке – закрыты.
В связи с этим различают 3 типа реакции устьичного аппарата на внешние условия среды:
1. Гидропассивные реакции – это закрытие устьиц в результате перенасыщения паренхимных клеток листа водой и механическое сдавливание устьиц, что закрывает устьичную щель. Это обычно наблюдается при обильном поливе дождеванием и может служить причиной подавления фотосинтеза.
2. Гидроактивная реакция закрывания или открывания устьиц – это движения, вызванные изменением в содержании воды в замыкающих клетках устьиц, что было рассмотрено выше.
3. Фотоактитвная реакция – связана с закрытием устьиц в темноте и открытием на свету... Согласно современным представлениям открывание устьиц индуцируется выходом Н+ из замыкающих клеток под действием света. Под действие света включается протонная помпа Н+- атефаза, которая выкачивает протоны из замыкающих клеток в клетки примыкающие. Поставщиком АТФ является процесс фотосинтеза. На смену протона в замыкающие клетки из прилегающих поступает ион К+ и для уравновешивания ион СL– и накапливаются в вакуолях повышая в них осмотический потенциал. С другой стороны, выход протона Н+ водорода, повышает рН среды и в щелочных условиях усиливается активность фермента ФЭПВК – карбокилазы ( фосфоэнол-карбоксилазы), которая ускоряет процесс карбоксилирования ФЭПВК (фосфоэнолпировиноградная кислота) и превращает её в малат (яблочная кислота) , тоже осмотические активное вещество. Ионы К+ и СL– и малат приводят к усилению поступление воды в вакуоль, повышению тургора и открытию устьиц. Это можно представить в виде следующей схемы:
Свет
 | |||
 | |||
Включение протонной помпы
 ( выход Н+ из замыкающих клеток)
( выход Н+ из замыкающих клеток)
Транспорт К+ и CL– Повышение рН
 в замыкающие клетки
в замыкающие клетки
 Образование малата
Образование малата



 Повышение осмотического потенциала замыкающих клеток
Повышение осмотического потенциала замыкающих клеток
Поступление воды в замыкающие клетки
Открытие устьиц
Фотосинтез также влияет на движение устьиц. При интенсивном образовании углеводов возрастает сосущая сила клеток и устьица открываются Состояние устьиц зависит ои от СО2. Если концентрация в предустьичном пространстве падает ниже 0,03%, тургор замыкающих клеток увеличивается и устьица открываются. Частично с этим связано утреннее открывание устьиц ( уменьшается количество СО2 в результате возрастания фотосинтеза) Ночью в межклетниках концентрация СО2 увеличивается вследствие дыхания при уменьшении или отсутствии фотосинтеза –поэтому устьица закриваються. У суккулентов со специфичным суточным ритмом обмена органических кислот устьица открываются ночью, когда парциальное давление СО2 в межклетниках уменьшается вследствие интенсивного образования малата и закрываются днем, когда при декарбоксилировании малата высвобождается СО2
. Однако суточный ход устьичных движений в зависимости от внешних условий может у разных растений отклоняться от обычного. При нормальном водоснабжении обычно устьица утром открываются, вечером закрываются. Однако, степень открытия устьиц в течение дня зависит от условий влагообеспечения. Мало воды в листьях – они закрываются, достаточно – открываются. Такой ход характерен для злаков. Но есть растения, у которых ритм этот меняется. Если днем устьица были закрыты в зависимости от сложившихся условий водообеспечения, то ночью они открываются (люцерна, клевер, горох). И есть третья группа растений, у которых всегда днем устьица закрыты, а ночью открыты. Это пустынные растения суккуленты.
Дата добавления: 2015-08-11; просмотров: 6061;