При первичной обработке сигналов
Выделение составляющих векторных величин. В измерительных полях биодатчиков рецепторы могут иметь параллельное или веерообразное направление осей чувствительности, что позволяет биодатчиком измерить составляющие векторных величин.
При веерообразной структуре измеряемый вектор будет по-разному проектироваться на измерительные оси (рис. 10): рецепторы, чьи измерительные оси перпендикулярны этому вектору, определяют его направление (их выходные сигналы равны нулю); рецепторы с измерительными осями, совпадающими по направлению с вектором, дают сигналы, соответствующие его модулю. Сравнение выходных сигналов рецепторов информационного поля в нейронных цепях биодатчиков позволяет выделить информацию как о величине, так и направлении вектора.
 Рис. 10. Информационное поле силового биодатчика.
Рис. 10. Информационное поле силового биодатчика.
|  Рис. 11. Схема нейронных сетей глаз.
Рис. 11. Схема нейронных сетей глаз.
|
Вследствие этого, разные виды животных, решающие различные задачи навигации и управления, имеют неодинаковые структуры информационных полей одноименных биодатчиков. Например, утрикулусы у птиц обладают наибольшей чувствительностью к боковым составляющим ускорения, у пресмыкающихся – к продольным, у большинства млекопитающих этот биодатчик имеет одинаковую чувствительность как к продольным, так и к боковым линейным ускорениям.
Усиление контраста сигналов. Использование множества рецепторов для измерения реакции чувствительного элемента на внешнее воздействие позволяет решить задачу усиления сигналов. Такая первичная обработка информации осуществляется во всех биодатчиках, однако наиболее важное значение она имеет для пространственных ориентаторов, получающих изображения картин местности.
Усиление контраста изображений осуществляется непосредственно в нейронных сетях сетчаток глаз (рис. 11). Суть этого процесса заключается в сравнении выходных сигналов расположенных рядом фоторецепторов и обнаружении между ними некоторого различия. Если интенсивности светового излучения, попадающего на соседние фоторецепторы, отличаются друг от друга, соответственно будут различными и сигналы на выходе фоторецепторов. В нейронных сетях с помощью разностных сигналов это различие выявляется и меньший сигнал еще больше уменьшается (тормозится), а более мощный сигнал – усиливается (возбуждается). В результате такой обработки информации выходные сигналы фоторецепторов приобретают еще больше различия.
Усиление контраста между сигналами соседних фоторецепторов позволяет уже на этом уровне обработки информации обнаруживать контрастные объекты, выделять границы изображений и с высоким быстродействием включать контуры автоматического управления. В вышестоящие отделы мозга информация направляется в более концентрированной форме для дальнейшей обработки и распознавания.
Измерение частотных характеристик сигналов. Подобный способ получения и использования избыточной информации используется в различных органах чувств, однако наиболее ярко он проявляется в зрительных органах животных и человека, что позволяет глазам видеть все многообразие красок.
В сетчатках глаз человека усилительные фоторецепторы имеют одинаковые по форме колоколообразные спектральные характеристики, но сдвинутые относительно друг друга по длине волны. Вследствие этого фоторецепторы разделяются на три вида, каждый из которых имеет максимальную чувствительность только к определенной части спектра длин волн электромагнитных излучений: коротковолновой (синий), средней (зеленой) и длинноволновой (красной). Избирательность восприятия длин волн излучений обусловлена различиями в свойствах пигмента во внутренних структурах фоторецепторов, который поглощает кванты света и вызывает соответствующую реакцию рецепторной клетки.

Рис. 12. Спектральные характеристики фоторецепторов глаза человека.
Совместная обработка выходных сигналов трех видов фоторецепторов позволяет сформировать цветовое ощущение.
Выходной сигнал одного фоторецептора как измерительного преобразователя является информационно недостаточным, так как зависит от интенсивности воспринимаемого излучения, его длины волны, причем неоднозначно: одно и то же значение выходного сигнала возбуждается излучениями разных длин волн. Поэтому любое количество одинаковых фоторецепторов не позволит зрительному анализатору выявить цвет изображения.
Использование двух фоторецепторов, имеющих сдвинутые по частоте спектральные характеристики, можно, позволяет исключить влияние интенсивности светового воздействия, но неопределенность частотных его характеристик сохраняется. И только наличие трех фоторецепторов, имеющих сдвинутые спектральные характеристики, позволяет достоверно определить и длину волны, и интенсивность светового сигнала. Это означает, что трех типов фоторецепторов вполне достаточно для полноценного приема информации о цветном изображении воспринимаемой картины.
Таким образом, триада фоторецепторов образует измерительную систему, обладающую информационной достаточностью для однозначного определения амплитудно-частотных характеристик светового воздействия путем измерения амплитуд выходных сигналов. При этом необходимо выполнение двух условий:
а) спектральные характеристики фоторецепторов должны охватывать весь диапазон измерений глаза;
б) собственные частоты фоторецепторов не должны совпадать между собой, а быть равномерно распределены в пределах диапазона измерений.
Алгоритм обработки такой информации в настоящее время не известен, однако можно предположить, что в нейронных структурах сетчатки осуществляют алгебраическое суммирование сигналов триады фоторецепторов с определенными весами. В создании цветового восприятия также принимают участие фоторецепторы фоновой области сетчатки, которая измеряет интенсивность и длину волны света, рассеянного внутренними прозрачными структурами глаза (нуль-цвет). Участие сигнала нуль-цвета значительно обогащает палитру зрительного восприятия, например, воспринимать белый и черный цвета.
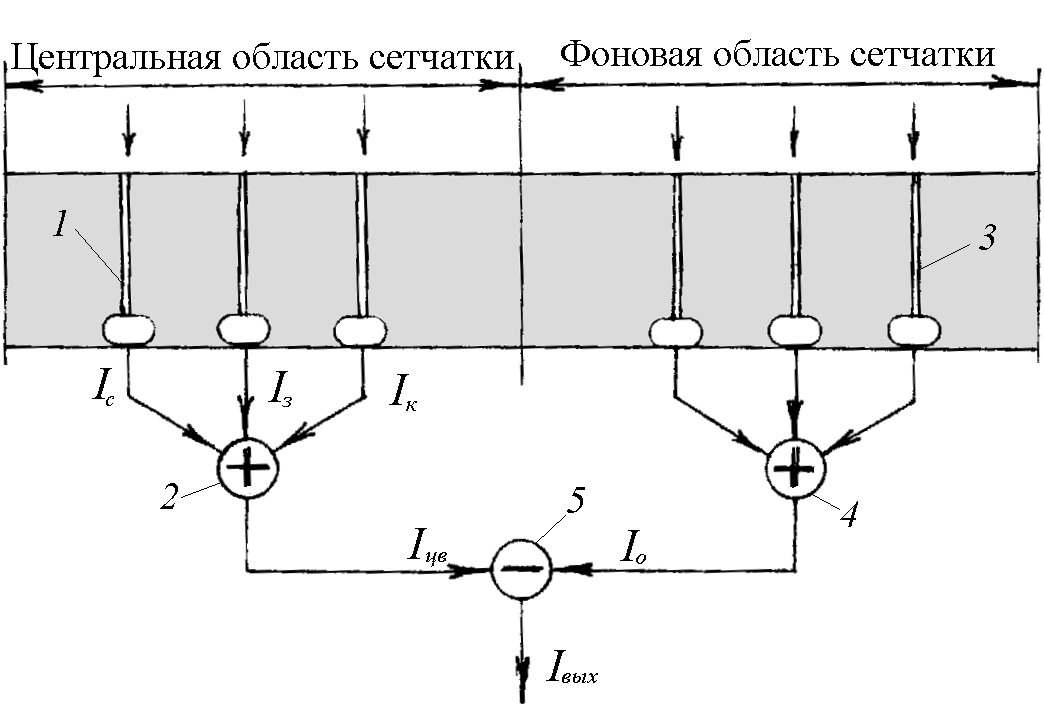
Рис. 13. Определение цветовой характеристики излучения.
Таким образом, в сетчатке глаза выполняется несколько операций по первичной обработке сигналов фоторецепторов: усиливается контраст, выделяются границы областей одинаковой яркости излучения и определяются цветовые характеристики излучения. Наличие такой информации дает возможность зрительному анализатору формировать изображение видимой картины. При этом в воспринимаемом глазом изображении он выделяет области, в которых отсутствует контраст, и окрашивает эти области в один цвет.
Компенсация динамических погрешностей. Высокая инерционность чувствительных элементов биодатчиков приводит к тому, что компенсация динамических погрешностей измерений является жизненно важной задачей живых существ. Решается эта задача непосредственно в их нейронных структурах в процессе первичной обработки сигналов.
Рассмотрим, каким образом обеспечивается высокое быстродействие биодатчиков на примере зрения человека. Информационное поле зрительного аппарата имеет неравномерное распределение фоторецепторов: центральная область сетчатки включает только усилительные ИП (колбочки), а периферическая – усилительные и адаптивные (палочки), причем последних в 150 раз больше, чем усилительных. Это означает, что основная функциональная роль в периферической области сетчатки принадлежит именно адаптивным фоторецепторам, способным выявлять изменения в наблюдаемой картине местности.
Чтобы понять, каким образом в результате первичной обработки информации обеспечивается компенсация динамического запаздывания сигналов в периферической области сетчатки глаза, сравним динамические свойства обеих измерительных структур сетчатки.
В центральной области сетчатки световые сигналы воспринимаются только одним типом фоторецепторов, передаточная функция которых соответствует инерционному звену. Участие множества усилительных фоторецепторов обеспечивает значительное увеличение коэффициента передачи сигнала. Суммарная передаточная функция измерительной структуры центральной области сетчатки также соответствует инерционному звену, вследствие чего постоянная времени зрительного восприятия в режимах распознавания объектов или слежения за движущимися целями составляет не менее 0,17с.
Световые сигналы, поступающие на периферическую область сетчатки, воспринимается двумя видами фоторецепторов: усилительными и адаптивными, которые имеют разные передаточные функции. При суммировании в определенной пропорции сигналов обоих видов фоторецепторов получается полная компенсация динамических погрешностей фоторецепторов в периферической области сетчатки глаза. Благодаря этому человек обладает высокой реакцией бокового зрения.
Таким образом, благодаря наличию адаптивных фоторецепторов в области периферического зрения не только выявляются изменения световых воздействий, но и обеспечивается компенсация инерционности чувствительных элементов глаз. Объединение выходных сигналов определенного количества палочек и колбочек в одном нейронном узле сетчатки создает форсирующее звено с характеристиками, необходимыми для компенсации динамических погрешностей канала измерения.
Дата добавления: 2015-08-11; просмотров: 1227;