Направленный рост аксонов
Направленный рост аксонов, как и адресная миграция клеток, осуществляется в рамках концепции “сигнал–ответ”. Эта концепция объясняет, как нейрон узнаёт свою область иннервации и находит своего клеточного партнёра, и как в развивающемся мозге многочисленные переплетающиеся отростки нейронов устанавливают связи с высокой точностью. Отросток нейрона — аксон — сразу и без ошибок находит свои мишени. Направленный рост аксонов осуществляет конус роста.
Конус роста — мобильная терминаль отростка нейрона, специализированная на удлинение и навигацию этого отростка, впервые детально описан Сантъяго Рамон-и-Кахалем. Конус имеет на конце булавовидное утолщение (ламеллоподия), от которого отходят тонкие пальцевидные отростки — филоподии. Они растут в различных направлениях и исследуют потенциальное пространство роста аксона. Согласно представлению С. Рамон-и-Кахаля о хемотропизме, рост аксонов происходит по градиенту концентрации специфических химических факторов, вырабатываемых в мишенях. Действительно, in vitro градиенты фактора роста нервов (NGF) и других веществ (например, ацетилхолина) влияют на направление роста аксонов. Наиболее разработано представление о меченых путях, которые образованы молекулярными метками (своего рода знаками навигационной обстановки), закономерно распределёнными в потенциальном пространстве роста аксонов. По мере роста пионерский аксон последовательно считывает одну за другой метки, расположенные в межклеточном пространстве или на поверхности клеток, и растёт в нужном направлении. Вслед за ним мигрируют отростки других аксонов, совокупность которых формирует тракты в ЦНС и нервы на периферии. Примером клеток, направляющих рост аксонов, могут служить временно живущие нейроны Кахаля–Ретциуса. Ключевой момент представления о заранее размеченных путях — узнавание — обеспечивают молекулы адгезии, которые встроены в плазмолемму ламеллоподии и филоподий и взаимодействуют с комплементарными молекулами (ламинин, фибронектин, коллаген, тенасцин и др.) во внеклеточном матриксе. Это обеспечивают фиксацию конуса роста на поверхности мишени в нужном месте и в нужное время.
Специфические сигналы из микроокружения позитивно регулируют, «разрешают» (хемоаттракция, хемоаттрактанты; от лат. attraho —— притягивать к себе) или, наоборот, негативно регулируют, «запрещают» (хеморепульсия, хеморепелленты, от лат. repellere —— отталкивать, отвергать) перемещение конуса роста в определённом направлении (рис. 8-10). Молекулы внеклеточного матрикса служат лигандами и взаимодействуют со своими рецепторами (интегринами) в аксолемме конуса роста (рис. 8-11, табл. 8-1). Конус роста аксона и направляющая его рост клетка могут взаимодействовать путём Ca2+-независимой адгезии с участием молекул L1 или NCAM (Neural Cell Adhesion Molecule), которые одновременно выступают в роли и лигандов, и рецепторов (гомофильное взаимодействие). Подобное гомофильное взаимодействие может осуществляться также путёмCa2+-зависимой адгезии при помощи кадгеринов. Секреторные (т.е. не фиксированные на поверхности, а выделяемые клеткой) хемоаттрактанты и хеморепелленты объединены в семейство нетрина/slit. Нетрин действует на клетку-мишень через рецептор DCC (Deleted in Colorectal Cancer), а молекулы slit действуют через рецептор Robo (Roundabout). Типичными репеллентами являются семафорины, которые могут секретироваться из клетки или быть связанными с её поверхностью. Рецепторы семафоринов (плексины и нейропилин) встроены в аксолемму конуса роста. Действие трансмембранных или мембрано-ассоциированных молекул из семейства эфринов осуществляется через рецепторные тирозинкиназы Eph.
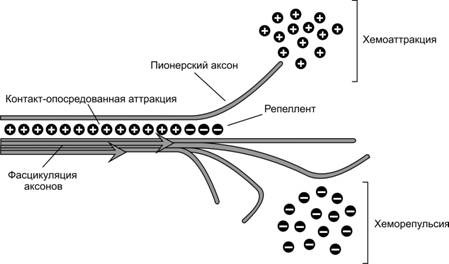
Рис. 8-10. Рост аксонов в заданном направлении контролируется распределением молекул аттрактантов и репеллентов.Конус роста пионерского аксона отрывается от группы аксонов и первым достигает области повышенной концентрации аттрактанта. Хеморепелленты отталкивают растущие аксоны. Рост аксонов компактным пучком (фасцикуляция) поддерживается молекулами адгезии в аксолемме (контакт-опосредованная аттракция). Если в определенном участке подобного пучка исчезают молекулы-аттрактанты и появляются молекулы-репелленты, то в этом участке происходит дефасцикуляция — аксоны «разбегаются» в разные стороны. [128]

Рис. 8-11. Основные семейства молекул, контролирующих направленный рост аксонов.Эти молекулы распределены в пространстве роста аксона и присутствуют во внеклеточном матриксе или связаны с мембраной направляющей рост аксона клетки (верхняя часть каждого рисунка). Эти лиганды узнаются рецепторами, встроенными в аксолемму конуса роста (нижняя часть каждого рисунка). Сигналы взаимодействия лиганд–рецептор передаются на внутриклеточные каскады, регулирующие сборку актиновых микрофиламентов в конусе роста. [128]
Таблица 8-1. Молекулы, контролирующие направленный рост аксона
| Семейство | Лиганды | Рецепторы | Характеристика |
| Молекул внеклеточного матрикса | Ламинин, тенасцины, коллаген, фибронектин, иммуноглобулины, аносмин‑1 (KAL), хондроитин/гепарансульфат протеогликаны, тромбоспондин‑1 | HomA, протеогликаны, интегрины | Многие из них важны для нейроонтогенеза. Все продолжают экспрессироваться в зрелом мозге |
| Семафоринов/коллапсинов | Sema 3A‑F, Sema 4A‑G, Sema 5A и B, Sema 6A‑C, Sema 7A | Плексин A, нейропилин | Хеморепелленты, вызывают коллапс конуса роста сенсорных аксонов in vivo. Некоторые разновидности могут выступать в роли аттрактантов. Могут быть интегральными мембранными белками или секретироваться |
| Иммуноглобулинов | N–CAM, PSA-N–CAM,L1,TAG-1/аксонин‑1,DM-Grasp, кадгерины | HetA, HomA | Интегральные и мембраносвязанные гликопротеины, участвуют в прикреплении аксонов к субстрату, фасцикуляции, направленном росте, в гомофильных и гетерофильных взаимодействиях |
| Миелин-ассоциированных ингибиторов | Миелин-ассоциированный гликопротеин (MAG), nogo | Неизвестны | Интегральные мембранные белки экспрессируются в зрелом миелине, ингибиторы роста аксонов in vitro и in vivo. Могут выступать в роли рецепторов для некоторых молекул внеклеточного матрикса |
| Рецепторов, связанных с тирозинкиназам | Ephrin A1-A5, Ephrin B1-B3 | Eph A1-A8 Eph B1-B6 | Лиганды и рецепторы, связаны с мембраной или являются трансмембранными белками, участвуют в процессах миграции клеток и роста аксонов в нейроонтогенезе |
| Нетринов | Нетрины 1 и 2 | DCC, нейрогенин,Unc‑5 | Важны для миграции нейробластов и направленного роста аксонов в развивающемся мозге. Связывание с рецептором DCC прослежено как при позитивном, так и негативном регулировании |
| Нейротрофических факторов | NGF, NT-3, BDNF, CNTF, GDNF | Trk A,B,C, p75, Lifrb, GFRa1 | Нейротропная и нейротрофичекая активность |
| Факторов роста | FGF, IGF, PDGF, VEGFa | FGFr1-4, flk-1, IGFr1, PDGFr, EGFr | Нейротрофическая активность, контроль пролиферации, дифференцировки и гибели клеток |
| Цитокинов | TGFb, LIF, TNF, EGF | TGFb тип I; TGFb,тип II; LIFrb, gp130, TNFr1-2 | Нейротрофичекая активность, поддержание дифференцировки нейронов |
| Нейромедиаторов | Ацетилхолин | Холинорецептор | Возможно, участвуют в синаптогенезе. Изменяют направление роста аксонов чувствительных нейронов |
| Slit | Slit–белки | Robo | Репелленты для различных аксонов и мигрирующих клеток, но и стимуляторы ветвления и удлинения аксонов |
Дата добавления: 2015-07-07; просмотров: 2858;