Дифференцировка остеобластов
Остеобласты и хондроциты происходят из общей плюрипотентной стволовой мезенхимной клетки (рис. 6-60, 6-61). Cbf1 (Core-binding factor -1) — один из важнейших транскрипционных факторов, регулирующих экспрессию многих остеоспецифических генов и их дифференцировку. В отсутствие хотя бы одного аллеля Cbf1 развивается ключично-черепная дисплазия. В зрелых остеобластах генами-мишенями транскрипционного фактораCbf1 служат гены остеокальцина, сиалопротеина кости, остеопонтина и субъединицы 1 коллагена. В свою очередь, экспрессию Cbf1 индуцируют морфогенетические белки кости (BMP).
Наряду с Cbf1, дифференцировку остеобластов регулирует транскрипционный фактор Osterix (Osx), или Sp7. Фактор транскрипции Sp7 связан с Cbf1 и не экспрессируется в отсутствие Cbf1. Osterix связывается с последовательностями Sp1 и EKLF, содержит 3 C2H2-типа «цинковые пальцы», относится к семейству белков Sp(Specificity protein).
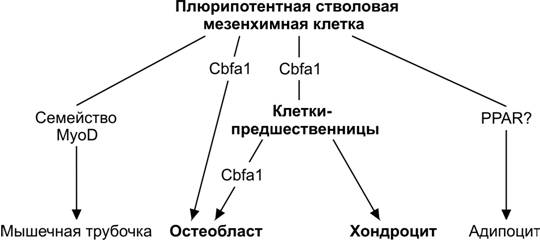
Рис. 6-60. Направления дифференцировки мезенхимной клетки.Остеобласты, хондроциты, мышечные трубочки и адипоциты происходят из общей плюрипотентной стволовой мезенхимной клетки. Каждый дифферон контролируется специфическим транскрипционным фактором. Мышечно-специфический фактор транскрипции, антиген миогенной дифференцировки MyoD (Myogenic Differentiation antigen) активирует мышечные гены и контролирует дифференцировку миобластов. -Рецептор пролифератора пероксисом (PPARγ) индуцирует дифференцировку адипоцитов. Транскрипционные факторы для хондрогенной дифференцировки пока не установлены. [139]
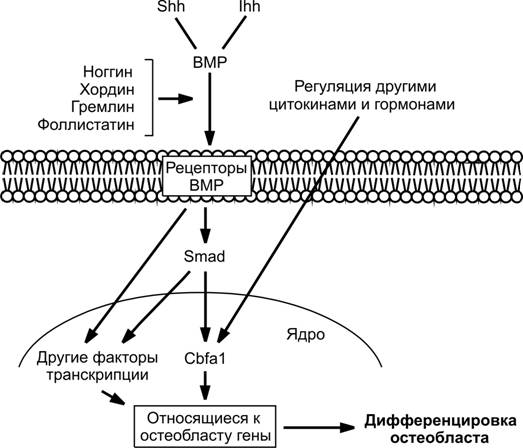
Рис. 6-61. Молекулярные пути дифференцировки остеобластов.Молекулярные факторы Sonic hedgehog (Shh) и Indian hedgehog (Ihh) действуют совместно сморфогенетическими белками кости(BMP). Ноггин и хордин подавляют эффекты BMP, конкурируя за рецепторыBMP, встроенные в клеточную мембрану (см. главу 3). BMP усиливают экспрессию Cbf1, что опосредуется транскрипционными факторами семейства Smad (Sma и MADD (Mother against decapentaplegic) — цитоплазматические посредники, которые активируются путем фосфорилирования рецептором ростовых факторов TGF. Внеклеточными лигандами для подобной активации служит не толькоTGF, но и другие члены одноименного надсемейства, а именно активины и BMP. Cbf1 поддерживает экспрессию генов остеогенной дифференцировки. Некоторые другие транскрипционные факторы, экспрессия которых усиливается в обход сигнального пути Cbf1 через молекулы Smad и рецепторный вход для BMP, также вовлечены в процесс дифференцировки остеобластов. [139]
Остеобласты начинают вырабатывать остеоид. Остеоид минерализуется, и дифференцирующиеся остеоциты оказываются замурованными в лакунах минерализованного костного матрикса. Сформировавшаяся незрелая грубоволокнистая костная ткань существует в форме трабекулы. Отдельные трабекулы, образовавшиеся в различных участках, растут и объединяются друг с другом. Наиболее толстые трабекулы (диаметром свыше 0,4 мм) содержат кровеносный сосуд, расположенный в центральном узком канале, выстланном остеогенными клетками. Поверхность трабекул покрывает слой остеобластов и остеогенных клеток. За счёт этого слоя на поверхности незрелой костной ткани происходит образование костных пластинок. Постепенно остеокласты разрушают первичную кость и на её месте путём аппозиционного роста формируются слои параллельных пластинок, образующих костные трабекулы из зрелой костной ткани. Анастомозирующая сеть костных трабекул формирует губчатое вещество. С утолщением трабекул и уменьшением полостей между ними, вплоть до их исчезновения, губчатое вещество может перестраиваться в компактное, состоящее из остеонов. Длина остеонов плоских костей довольно мала по сравнению с остеонами длинных трубчатых костей. В плоских костях губчатое вещество сохраняется в виде очень тонкого среднего слоя — diploё.
Дата добавления: 2015-07-07; просмотров: 2577;