Под ТОРМОЖЕНИЕМ понимают самостоятельный нервный активный процесс, вызываемый возбуждением, который проявляется в угнетении или полном выключении другого возбуждения
Впервые возможность влияния на рефлекторные процессы без воздействия на структуры рефлекторной дуги была обнаружена И.М. Сеченовым. В 1863 году И.М. Сеченовым было открыто ЦЕНТРАЛЬНОЕ ТОРМОЖЕНИЕ (СЕЧЕНОВСКОЕ). И.М. Сеченову удалось установить, что сверхпороговое возбуждение центров среднего мозга приводит к торможению периферических, спинальных рефлексов.
Различают первичное и вторичное торможение. Первичное торможение обусловлено нервными импульсами, возникающими в особых тормозных нейронах.
В центральной нервной системе тормозные нейроны есть в спинном мозге, в головном мозге (меньшее количество) и в коре головного мозга (большинство). В спинном мозге 2 вида тормозных нейронов (это тормозные вставочные нейроны):
клетки Реншоу - не обладают фоновой активностью и в покое не генерируют нервных импульсов. Они возбуждаются под действием: импульсов от афферентных нейронов, от эфферентных нейронов (альфа-мотонейронов спинного мозга), импульсов от вышележащих отделов головного мозга;
клетки Уилсона - обладают постоянной фоновой активностью, даже без раздражения (в покое) они генерируют нервные импульсы - постоянно тормозят активность альфа-мотонейронов спинного мозга.
В головном мозге существуют отдельные клетки Реншоу и клетки Пуркинье, грушевидные нейроны мозжечка - они оказывают торможение внутри мозжечка, ядер среднего и продолговатого мозга, тем самым обеспечивается правильное распределение мышечного тонуса.
В коре головного мозга 4 вида тормозных клеток:
1. большие корзинчатые нейроны - 3, 4, 5 слои коры головного мозга, их аксоны сильно ветвятся и образуют сплетения на площади около 500 мкм. Они тормозят активность нейронов 3, 4, 5 слоев;
2. малые корзинчатые клетки нейроны - 2, 3 слои коры - их аксоны ветвятся на меньшей площади, около 50 мкм, и тормозят 2 и 3 слои;
3. нейроны с кистеобразным аксоном - 1 слой коры, образует аксон, на конце которого разветвление в виде кисти; тормозит клетки 1-го слоя;
4. канделяброобразные нейроны - на границе между 2 и 3 слоями, вниз от них идет аксон и дает несколько ответвлений вверх, тормозят активность всех слоев.
Первичное торможение осуществляется за счет выделения тормозного медиатора на окончаниях нервных клеток (ГАМК - гаммааминомаслянная кислота, глицин, таурин, серотонин и другие). Эти вещества вызывают гиперполяризацию постсинаптической мембраны и, как следствие, тормозной постсинаптический потенциал.
Вспомним результат выделения медиатора пресинаптической мембраной: на постсинаптической мембране происходит деполяризация и возникает локальный ответ. Тормозные нейроны синтезируют другие медиаторы - тормозные. Эти медиаторы, взаимодействуя со своими рецепторами, открывают другие каналы, например калиевые или хлорные. Что же происходит в этом случае с постсинаптической мембраной?

Тормозной медиатор, как и возбуждающий, активирует рецептор управляемые каналы, однако не натриевые, а калиевые. Эта активация вызывает увеличение тока ионов калия. Если вспомнить о том, что ионы калия движутся по градиенту концентрации из клетки, то становится ясно, что при активации калиевых каналов происходит гиперполяризация мембраны и снижение возбудимости нейрона. Изменение потенциала мембраны при гиперполяризации называется тормозной постсинаптический потенциал (ТПСП). Во многих тормозных синапсах аналогичный эффект имеет активация каналов для хлора, увеличивающая его транспорт внутрь клетки. Результатом ТПСП является удаление МПП от критического уровня деполяризации возбуждение становится либо вовсе невозможным, либо для возбуждения требуется значительно большая сила раздражителя. Нервные импульсы, возникающие при возбуждении тормозных нейронов, вызывают гиперполяризацию постсинаптической мембраны и тормозной постсинаптический потенциал (ТПСП), это наиболее распространенный тип торможения в нервной системе, который называется ПОСТСИНАПТИЧЕСКИМ ТОРМОЖЕНИЕМ. Постсинаптическое торможение является первичным торможением – самостоятельный процесс, вызванный возбуждением тормозных нейронов.

Рисунок 25. Суммация потенциалов на мембране нейрона
Центральные синапсы отличаются тем, что медиаторы вызывают появление на постсинаптической мембране только локального ответа, (ВПСП или ТПСП). ВПСП обладает всеми свойствами ЛО - зависит от силы раздражителя (количества выделившегося медиатора), суммируется и не передается по аксону.
Амплитуда одиночного ВПСП довольно мала - несколько мВ, поэтому для уменьшения заряда мембраны до критического уровня необходима либо одновременная активация нескольких возбуждающих синапсов - ПРОСТРАНСТВЕННАЯ СУММАЦИЯ, либо повышение частоты поступающих импульсов - ВРЕМЕННАЯ СУММАЦИЯ. Если одновременно с ВПСП на мембране нейрона формируется и ТПСП, то эти изменения мембранного потенциала суммируются, следовательно, если величины ВПСП и ТПСП примерно одинаковы, то изменения мембранного потенциала нейрона, на котором суммируются воздействия, не произойдет (рис 25,26).
На рис.25 представлена схема регистрации мембранного потенциала возбуждающего и тормозного нейронов, которые одновременно воздействуют на один нейрон. И возбуждающий, и тормозной нейрон возбуждены – регистрируется ПД. Следовательно, одновременно выделяются и возбуждающий, и тормозной медиаторы. На мембране тестируемого нейрона одновременно возникают ВПСП и ТПСП примерно одинаковой амплитуды, в результате суммации мембранный потенциал остается неизменным.

Рисунок 26. Виды суммации на нейроне
Существует еще один вариант первичного торможения в ЦНС: ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ. Такое торможение возникает в том случае, когда нервный импульс или просто не достигает синаптического окончания аксона, или снижается его частота. Этот вид торможения осуществляется благодаря аксоаксональным синапсам, образованным терминалями аксонов тормозных интернейронов и аксонами возбуждающих нейронов (рис.26, справа). В этих синапсах торможение определяется действием ГАМК, открывающей хлорные каналы на пресинаптических мембранах (мембрана возбуждающего нейрона). В результате на постсинаптической мембране тормозного синапса возникает явление стойкой деполяризации. Деполяризация мембраны аксона ведет к инактивации натриевой проводимости. Возникает блокирование проведения нервного импульса, что уменьшает частоту, или полностью угнетает возбуждающий потенциал, идущий к клетке по возбуждающим нервным окончаниям - блок проведения возбуждения.
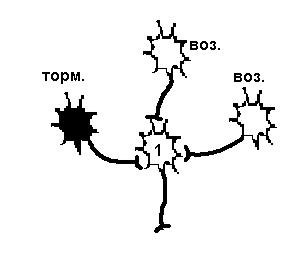
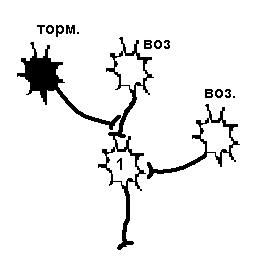
| Постсинаптическое торможение | Пресинаптическое торможение |
| 
|
| Возбудимость нейрона 1 снижена в результате гиперполяризации мембраны, вызванной действием тормозного нейрона, поэтому два возбуждающих нейрона не могут вызвать ПД. | Возбудимость нейрона 1 не изменена, но возбуждающий импульс от одного из возбуждающих нейронов не доходит, а один возбуждающий не может вызвать ПД. |
Рисунок 26. Первичное торможение
Вторичное торможение возникает в обычных возбудимых структурах и связано с процессом возбуждения. Виды вторичного Запредельное торможение - возникает в нейронах центральной нервной системы в том случае, когда поток информации к телу нейрона выше его работоспособности. Развивается резкое снижение возбудимости нейрона. Парабиотическое торможение - возникает при действии сильных и длительно действующих раздражителей (парабиоз в тканях). Парабиоз - явление пограничного состояния между гибелью и жизнью ткани (резко падают все свойства ткани, основное свойство - фазное изменение лабильности). Если парабиотический фактор продолжает действовать, ткань гибнет. Пессимальное торможение - возникает в синапсах центральной нервной системы при действии сильных и частых раздражителей. Торможение вслед за возбуждением - угнетение нейронов после возбуждения. Результат того, что вслед за пиком потенциала действия возникает период следовой гиперполяризации, который характеризуется снижением возбудимости.
Дата добавления: 2015-07-07; просмотров: 4262;