Общие пути обмена аминокислот.
Пути распада аминокислот до конечных продуктов можно разделить на 3 группы:
1. Пути распада, связанные с превращением NH2-групп.
2. Декарбоксилирование 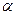 -СООН групп.
-СООН групп.
3. Превращения углеродного скелета аминокислот.
Превращение -аминогрупп аминокислот.
В тканях организма происходит отщепление аминогрупп с образованием аммиака. Этот процесс называется дезаминированием. Возможны 4 типа дезаминирования:
1. Восстановительное
R-CH-COOH 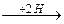 R-CH2-COOH + NH3
R-CH2-COOH + NH3
|
NH2
2. Гидролитическое
R-CH-COOH 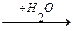 R-CH-COOH + NH3
R-CH-COOH + NH3
| |
NH2 OH
3. Внутримолекулярное
R-CH2-CH-COOH  R-CH=CH-COOH + NH3
R-CH=CH-COOH + NH3
|
NH2
4. Окислительное
R-CH-COOH 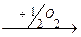 R-C-COOH + NH3
R-C-COOH + NH3
| ||
NH2 O
Окислительное дезаминирование бывает 2 видов: прямое и непрямое (трансдезаминирование).
Прямое окислительное дезаминирование осуществляется оксидазами. В качестве кофермента содержат ФМН или ФАД. Продуктами
реакции являются кетокислоты и аммиак.
Трансдезаминирование – основной путь дезаминирования аминокислот. Трансдезаминирование проходит в 2 этапа.
Первый – трансаминирование – перенос аминогруппы с любой аминокислоты на -кетокислоту без промежуточного образования аммиака;
Второй – собственно окислительное дезаминирование.
В результате первого этапа аминогруппы «собираются» в составе глутаминовой кислоты. Второй этап связан с окислительным дезаминированием глутаминовой кислоты.
Трансаминирование аминокислот было открыто советскими учеными Браунштейном и Крицман (1937г).
R R1 R R1
| | | |
HC-NH2 + C=O  C=O + HC-NH2
C=O + HC-NH2
| | | |
COOH COOH COOH COOH
Реакция трансаминирования обратима, она катализируется ферментами – аминотрансферазами.
Акцептором аминогрупп в реакциях трансаминирования являются три - кетокислоты: пируват, оксалоацетат, 2-оксоглутарат. Наиболее часто акцептором NH2-групп служит 2-оксоглутарат ( -кетоглутарат), реакция приводит к образованию глутаминовой кислоты:
1. СН3 COOH CH3 COOH
| | АЛТ | |
НСNH2 + CH2 C=O + (CH2)2
| | | |
COOH CH2 COOH CHNH2
| |
C=O COOH
| глутамат
COOH
Аминотрансферазы содержат в качестве кофермента производные пиридоксина (витамин В6) – пиридоксаль-фосфат и пиридоксамин-фосфат.
(Механизм реакции трансаминирования с участием пиридоксальфосфата – учебник.)
2. Окислительное дезаминирование глутаминовой кислоты.
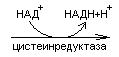
Биологический смысл реакций трансаминирования состоит в том, чтобы собрать аминогрупы всех распадающихся аминокислот в составе одной аминокислоты – глутаминовой. Глутаминовая кислота поступает в митохондрии клеток, где происходит второй этап трансдезаминирования – собственно дезаминирование глутаминовой кислоты. Реакция катализируется глутаматдегидрогеназой, которая в качестве кофермента содержит НАД+ или НАДФ+.
COOH СООН
| |
 (CH2)2
(CH2)2 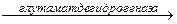 (CH2)2
(CH2)2
| |
CHNH2 НАД НАДН+Н+ C=O
| |
COOH COOH
Клиническое значение определения активности трансаминаз.
Для клинических целей определяют активность АлТ и АсТ, которые катализируют следующие реакции:
Аспартат + -кетоглутарат оксалоацетат + глутамат
Аланин + -кетоглутарат пируват + глутамат
В сыворотке крови здоровых людей активность этих трансаминаз ниже, чем в органах. При поражении органов наблюдается выход трансаминаз из очага поражения в кровь. Так, при инфаркте миокарда уровень АсТ сыворотки крови уже через 3-5 ч после наступления инфаркта повышается в 20-30 раз. При гепатитах повышается более умеренное и затяжное.
Декарбоксилирование аминокислот.
Процесс отщепление карбоксильной группы в виде СО2 называется декарбоксилированием и приводит к образованию биогенных аминов, которые оказывают фармакологическое действие на физиологические функции человека.
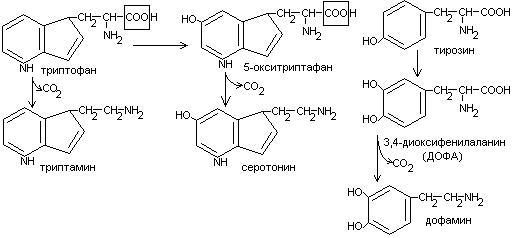
Серотонин обладает сосудосуживающим действием, участвует в регуляции артериального давления, t тела, дыхания, медиатор нервных процессов.
Дофамин- предшественник катехоламинов.
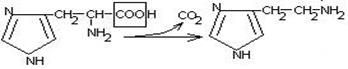
гистидин гистамин
Гистамин обладает сосудорасширяющим действием. Он образуется в области воспаления, участвует в развитии аллергических реакций.
НООС-(СН2)2-СН-СООН  СН2-СН2-СН2-СООН
СН2-СН2-СН2-СООН
| |
глутамат NH2 NH2  -аминомасляная кислота (ГАМК)
-аминомасляная кислота (ГАМК)
ГАМК является тормозным медиатором. В лечебной практике используется при лечении эпилепсии (резкое сокращение частоты припадков).
Орнитин декарбоксилируясь дает диамин – путресцин, а лизин – кадаверин.
СН2-СН2-СН2-СН-СООН СН2-СН2-СН2-СН2-NH2;
| | |
NH2 NH2 NH2
CH2-CH2-CH2-CH2-CH-COOH СН2-СН2-СН2-СН2-CH2-NH2
| | |
NH2 NH2 NH2
В организме биогенные амины подвергаются реакции окислительного дезаминирования с образованием альдегидов и аммиака. Процесс осуществляется при участии моноаминооксидаз.
Схематически механизм трансдезаминирования можно представит так:

Пути превращения аминокислот в печени.
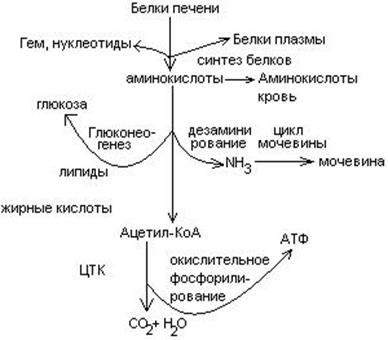
Включение углеродных скелетов аминокислот в цикл лимонной кислоты.
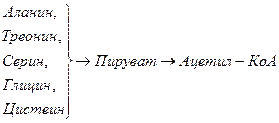
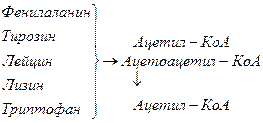
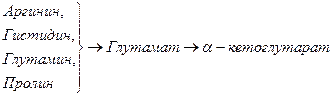
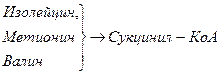
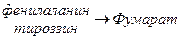
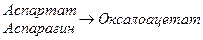
Синтез аминокислот
Заменимые аминокислоты образуются в печени из глутаминовой кислоты. Источником всех атомов углерода (а также азота аминогрупп) пролина, орнитина и аргинина является глутамат.
Источником углерода аланина, серина и глицина служат трехуглеродные промежуточные продукты гликолиза, а источником азота их аминогрупп – глутамат; аспартат получает углерод от оксалоацетата, а азот от глутамата.
-кетоглутарат + NH3 + НАДН (или НАДФН)  Глутамат + НАД+ (или НАДФ+)
Глутамат + НАД+ (или НАДФ+)
Глутамат + -кетокислота -кетоглутарат + аминокислота
-кетокислота + NH3 + НАДН (НАДФН) аминокислота + НАД+ (НАДФ+)
Восстаниовительное аминирование в печени и других тканях обеспечивает сохраниение и повторное использование аммиака. Большая часть аминокислот синтезируется в результате переноса аминогрупп от других аминокислот без превращения в NH3.
Трансаминирование = переаминирование = аминирование -кетоглутарата.
Мак-Мюррей, стр.276
Трансаминированию в печени не подвергаются аминокислоты с разветвленной цепью – лейцин, изолейцин, валин.
Эти аминокислоты вступают в реакции трансаминироания в почечной и мышечной тканях.
Лизин и треонин не вступают в реакцию трансаминирования.
Азотистый обмен
В организме человека ежесуточно распадается на аминокислоты около 400 г беков и столько их синтезируется.
Основная функция белков пластическая. Белки могут выполнять энергетическую функцию при избыточном их поступлении с пищей или в экстремальных ситуациях, когда белки тела подвергаются усиленному распаду. Этот процесс наблюдается при голодании или патологии (при сахарном диабете). При сгорании 1 г белков высвобождается 16,8 кДж.
Под термином «резервные» белки понимают не особые отложения белков, а легкомобилизуемые при необходимости тканевые белки, которые после гидролиза под действием тканевых протеиназ дают аминокислоты.
Факторы определяющие состояние белкового обмена.
1. Возраст (в детском возрасте, при активной мышечной работе, беременности), состояние организма – резко повышается потребность в белках.
2. Характер питания (количественный и качественный состав пищи). При недостаточном поступлении белков с пищей происходит распад белков ряда тканей (печени, плазмы, крови). Введение с пищей повышенных количеств белка не оказывает заметного влияния на состояние белкового обмена, если избыточное поступление белка не превышает 2-3 раза. Качественный состав белков важен, т.к. отсутствие или недостаток хотя бы одной какой-либо незаменимой аминокислоты нарушает процесс биосинтеза белков в организме. Синтез белка подчиняется закону «все или ничего» и осуществляется при условии наличия в клетке полного набора 20 аминокислот.
3. Обмен белков связан с обеспеченностью организма витаминами В1, В2, В6, РР и др.
4. Секреция гормонов (СТГ, гормоны щитовидной железы, андрогены, эстрогены).
Для оценки состояния обмена белков используется понятие азотистый баланс.
Азотистый баланс – разница между введением с пищей азота и выведением его в виде конечных продуктов азотистого обмена.
Различают 3 вида азотистого баланса:
1. Положительный – количество выводимого из организма азота меньше количества азота, вводимого с пищей. Азот остается в организме и расходуется на синтез белков. Характерен для растушего организма и во время беременности.
2. Отрицательный – количество выделяемого азота превышает количество азота, поступающего в течение суток. Встречается при голодании, белковой недостаточности, тяжелых заболеваниях, когда происходит интенсивный распад белков тела. Отрицательный азотистый баланс наблюдается у пожилых людей.
3. Азотистое равновесие – количество азота выводимого из организма, равно количеству получаемого с пищей азота. Характерно для здорового взрослого человека.
Биологическая ценность белков.
Определяется следующими критериями:
1. Чем ближе аминокислотный состав пищевого белка к аминокислотному составу беков тела, тем выше его биологическая ценность.
2. В составе белка должны быть все незамеимые аминокислоты: вал, илей, лей, лиз, мет, тре, три, фен. Исключение какой-либо незаменимой аминокислоты из пищи сопровождается развитием отрицательного азотистого баланса.
3. В составе белка установлены пропорциональные величины незаменимых аминокислот, относительно триптофана, принятого за единицу, лиз-5, лей-4, вал-3,5.
Недостаток в пище одной незаменимой аминокислоты ведет к неполному усвоению других аминокислот.
Для человека наиболее ценны белки молока, яиц, мяса, т.к. их аминокислотный состав ближе к аминокислотному составу органов и тканей человека.
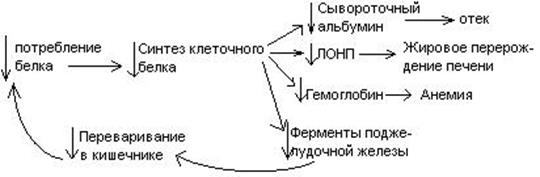
В результате недостаточного поступления белков в организм возникает синдром кваширхор. Поражает детей в период отнятия от груди. Нарушение норамального синтеза белка в печени вызывает снижение содержания сыворотного альбумина, что приводит к отеку, снижение содержания ЛОНП приводит к развитию жирового поражения печени. Снижен синтез гемоглобина, в результате развивается анемия. Резко нарушается функция кишечника, т.к. из недостатка аминокислот страдает синтез ферментов поджелудочной железы. Смерть наступает от диареи, острых инфекций и хронических заболеваний печени.
Переваривание и всасивание
Переваривание белков происходит в 3 этапа:
1. в желудке;
2. в тонком кишечнике;
3. в клетках слизистой оболочки тонкого кишечника.
Расщепление белков происходит при участии нескольких групп ферментов:
1. Экзопептидазы – катализирует разрыв концевой пептидной связи с образованием одной какой-либо аминокислоты.
2. Эндопептидазы – гидролизуют пептидные связи внутри полипептидной цепи.
К эндопептидазам относятся следующие ферменты: пепсин, гастрин, трипсин, химотрипсин, эластаза.
К экзопептидазам относятся: карбоксипептидазы, аминопептидазы, дипептидазы.
Экзопептизазы участвуют в переваривании белков в тонком кишечнике.
В результате расщепления образуются свободные аминокислоты, которые затем подвергаются всасыванию. Аминокислоты всасываются свободно с ионами натрия. Некоторые аминокислоты обладают способностью конкурентно тормозить всасывание других аминокислот:
Лизин тормозит всасывание аргинина. Это позволяет считать, что существует общая переносящая система.
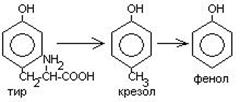
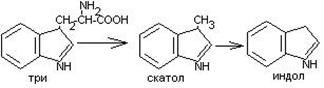
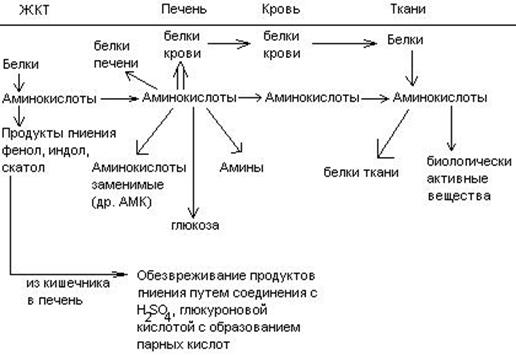
Часть аминокислот в кишечнике под действием микрофлоры подвергается гниению. В результате гниения аминокислот в кишечнике образуются ядовитые продукты распада – фенол, индол, крезол, скатол, сероводород. Распад цистеина, цистина, метионина приводит к образованию сероводорода, метилмеркаптана. Диаминокислоты – арнитин и лизин подвергаются декарбоксилированию с образованием путресцина и кадаверина.
Микробные ферменты кишечника превращают тирозин крезол фенол
триптофан скатол индол.
После всасывания эти продукты через воротную вену попадают в печень, где обезвреживаются путем связывания с серной или глюкуроновой кислотами с образованием парных кислот, которые выделяются с мочей.
Аминокислоты как лекараственные препараты.
Препараты гидролизатов белков – используются для перентерального питания.
К ним относятся гидролизин, гидролизат казеина, аминопептид, церебролизин, аминокровин, фиброносол.
Применяют после операций на ЖКТ, у больных с нарушениями переваривания белков и аминокислот, при тяжелых ожегах.
Препараты отдельных аминокислот.
Метион – липотропный фактор, а также для лечения белковой недостаточности при хронических заболеваниях.
Цистеин – при нарушениях обмена серосодержащих белков (хрусталик, роговица глаза, коллаген), при отравлениях солями тяжелых Ме, которые связываются аминокислотами.
Глутаминовая кислота и аспарагиновая кислота (калиевые и магниевые соли – препарат панангин, аспаркам) используются для обезвреживания аммиака.
Пути обезвреживания аммиака.
Уровень аммиака в норме в крови не превышает 60 мкМоль/литр. Для кроликов концентрация аммиака 3 мМоль/литр является летальной.
В организме существует 4 пути обезвреживания аммиака.
1. 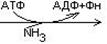 Образование глутамина – процесс идев в мозге, сетчатке, почках, печени и мышцах.
Образование глутамина – процесс идев в мозге, сетчатке, почках, печени и мышцах.
НООС-СН-СН2-СН2-СООН НООС-СН-(СН2)2-СONH2
| |
NH2 NH2
Глутамин выделяется с мочой.
2. Восстановительное аминирование -кетоглутаровой кислоты
-кетоглутарат+NH3 + НАДН глутамат + НАД+
Эта реакция обратная окислительному дезаминированию глутяминовой кислоты. Дальше глутамат связывает еще одну молекулу NH3 и дают глутамин. Глу+NH3 Глн.
Глутамин используется почками как резервный источник аммиака, необходимый для нейтрализации кислых продуктов обмена при ацидозе.
3. Образование аммонийных солей.
Аммиак в почках акцептирует протон кислоты, образуя аммонийную соль. Одновременно этот процесс обеспечивает сбережение организмом ионов Na+, которые в отсутствие ионов аммония выводились бы с анионами кислот.
NH3+Н+Cl- NH4Cl
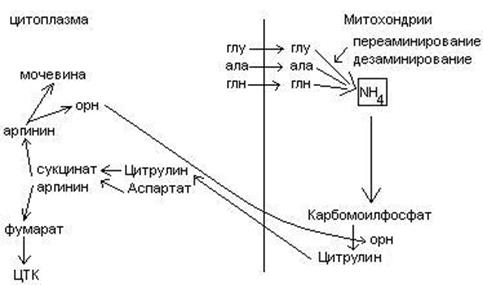
4. Биосинтез мочевины.
Мочевина – главный конечный продукт обмена азота в организме. С мочой за сутки выводится 25-30 г мочевины. Синтез мочевины идет в печени. Превращение аммиака в мочевину осуществляется в форме цикла. Цикл мочевины открыли Ганс Кребс и Курт Хенселайт (1932г).
Пернвая аминогруппа, поступающая в цикл мочевины, получается в виде свободного аммиака при окислительном дезаминировании глутамата в митохондриях клеток печени. Эта рекция катализируется глутаматдегидрогеназой, для действия которой требуется НАД+
Глутамат+НАД++Н2О -кетоглутарат + 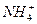 +НАДН+Н+.
+НАДН+Н+.
Аммиак соединяется с двуокисью углерода с образованием карбомоилфосфата. Реакция требует затраты АТФ. Катализирует эту реакцию карбомоилфосфатсинтетаза. На следующей стадии карбомоилфосфат взаимодействует с орнитином с образованием цитрулина. Образовавшийся цитрулин переходит из митохондрий в цитозоль клеток печени. Цитрулин взаимодействует с аспартатом в присутствии АТФ с образованием аргининсукцината. Эта реакция катализируется аргининсукцинатсинтетазой. На следующей стадии аргининсукцинат расщепляется с образованием аргинина и фумарата. Фумарат возвращается в пул промежуточных продуктов цикла лимонной кислоты. На последней стадии аргинин под действием аргиназы расщепляется на мочевину и орнитин.
Регенерированный орнитин вновь поступает в митохондрии.
Мочевина из клеток печени поступает в кровь и через почки выводится с мочой.
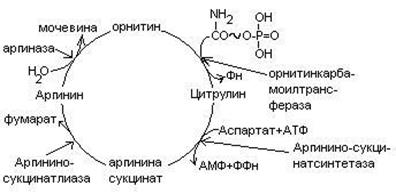
Повышение концентрации аммиака в крови может вызвать повторяющуюся рвоту, возбуждение, припадки с потерей сознания (печеночная кома) и судорогами. При хронической врожденной гипераммониемии наблюдается отставание умственного развития. Наиболее частая причина гипераммониемии – нарушение орнитинового цикла. В орнитиновом цикле участвует 5 ферментов и имеет место 5 типов наследственных болезней. Люди с такими дефектами не переносят пищи, богатой белком. Детей с таким нарушением лечат введением -кетоаналогов аминокислот. -кетоаналоги незаменимых аминокислот могут под действием трансаминаз присоединять аминогруппы от имеющихся заменимых аминокислот. Это предотвращает возможность попадания аммиака в кровь.
Почему высокое содержание NH4+ токсично? Ион аммония сдвигает равновесие реакции, катализируемой глутаматдегидрогеназой, в сторону образования глутамата, а это приводит к истощению -кетоглутарата. Истощение -оксоглутамата, промежуточного продукта ЦТК, приводит к уменьшению скорости образования АТФ.
-оксоглутарат 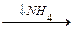 глутамат глутамин
глутамат глутамин
глутаматдегидрогеназа глутаминсинтетаза
Энергетическая цена синтеза мочевины
На синтез одной молекулы мочевины расходуется 4 высокоэнергетические фосфатные группы. Две молекулы АТФ требуются для образования аргининосукцината. Но в последней реакции АТФ подвергается пирофосфат, который гидролизуется с образованием 2 молекул ортофосфата. Поэтому на синтез мочевины расходуется 4 молекулы АТФ.
Синтез заменимых аминокислот
1. Аминирование
СООН COOH
| НАДФН2+NH4+ НАДФ |
 (CH2)2 (CH2)2 про
(CH2)2 (CH2)2 про
| | глн
C=O CHNH2
| |
COOH COOH
2. Переаминирование
Глу + пир ала + -КГ
Глу + ЩУК асп + -КГ
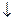
асн
3. Глю сер гли
Токсическое действие аммиака.
+ -КГ Глу  Глн
Глн
ЦТК  ЦТК АТФ в мозге Кома
ЦТК АТФ в мозге Кома
Способ лечения:
1. Перевод на малобелковую пищу
2. Замена заменимых аминокислот на кетокислоты (незам)
Гепатоцит
Индивидуальные пути обмена аминокислот.
Обмен глицина и серина.
Гли и сер относятся к заменимым, глюкопластичным аминокислотам.
- Глицин, много содержится в белках, не имеет радикала, оптически неактивна, заменимая, глюкогенная NН2-СН2-СООН
Используется для синтеза креатина в почках
Почки
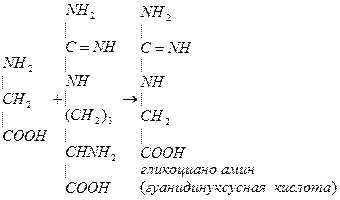
 Печень
Печень 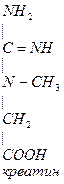
Мышцы требуют Е (кратковременная интенсивная работа). Должен быть запас Е-креатинфосфат.
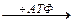
Может образовываться в других клетках. Это средство транспорта Е внутри клетки.
Фосфокреатин используется мышцами для кратковременной работы.
Продукт распада креатина креатин
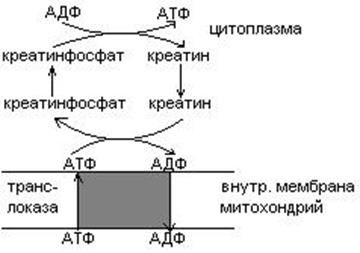
Креатинфосфат легко проходит в цитоплазму, он менее полярен, чем АТФ. Креатинфосфат отдает Фн на АДФ, а креатин возвращается в митохондрии. Такая транспортная функция креатинфосфата характерна для всех клеток.
- Образование
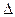 -аминолевулиновой кислоты.
-аминолевулиновой кислоты. - Синтез пуриновых оснований. Глицин является остовом – это скелет.
- Синтез вторичных желчных кислот. Желчные кислоты участвуют в переваривании жиров, холестеридов. Желчные кислоты активируются глицином (холевая – гликохолевая).
Глицин – нейромедиаторная АМК – тормозной медиатор нервной системы (30 минут). Глицин принимает участие в синтезе белков, пуриновых нуклеотидов, гема, парных желчных кислот, кератина, глутатиона.
 Глутатион Глутатион
| Сер |  Тканевые белки Тканевые белки
|
|  Глюкоза Глюкоза
| |
   Муравьиная кислота Муравьиная кислота
| Гли |     Липиды Липиды
|
| Гиппуровая кислота |
| Гем |
| Креатин | Тре | Пурины (ДНК, РНК) |
| Желчные кислоты |
Глицин участвует в образовании гема:
СООН СН2-NH2 HSKoA COOH
 | | B6 |
| | B6 |
 СН2 + COOH CH2 + CO2
СН2 + COOH CH2 + CO2
|  -аминолевули- |
-аминолевули- |
СН2 натсинтаза CH2
| |
COSKoA C=O
|
CH2-NH2
-аминолевулиновая кислота
В качестве кофермента -аминолевулинансинтаза содержит витамин В6. -аминолевулинсинтаза – аллостерический, ключевой фермент синтеза гема. Ингибируется активность фермента по принципу обратной отрицательной связи – гемом.
Глицин принимает участие в синтезе креатина. Креатин обеспечивает работающую мышцу АТФ. Синтез креатина идет в почках и печени. В почках образуется гуанидинуксусная кислота:
NH2 NH2 NH2 NH2
| | | |
C=NH + CH2 (CH2)3 + C=NH
| | | |
NH COOH CH-NH2 NH
| глицин | |
(CH2)3 COOH CH2
| орнитин |
CHNH2 COOH
| гуанидинуксусная
COOH кислота
Аргинин
Гуанидинацетат с кровотоком поступает в печень, где в результате реакции трансметилирования дает креатин:
NH2 NH2 NH~PO3H2
| | |
 C=NH CH3 C=NH АТФ C=NH
C=NH CH3 C=NH АТФ C=NH
 | | |
| | |
NH N-CH3 N-CH3
| | |
CH2 CH2 CH2
| | |
COOH COOH COOH
Креатин Креатинфосфат
Источником метильного радикала является метионин.
Серин – заменимая аминокислота, углеродная часть которой образуется из глюкозы:


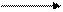 Глю
Глю  3-ФГК
3-ФГК 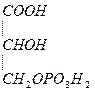 НАД НАДН
НАД НАДН 

 глутамат -кетоглутарат
глутамат -кетоглутарат 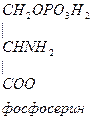
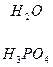
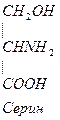
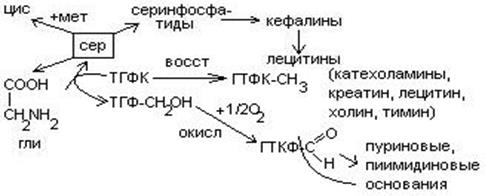
Серин – содержит ОН–группу, заменимая, глюкогенная. Является источником одноуглеродных фрагментов, которые идут на синтез БАВ (гормоны, медиаторы).
Серин является донатором одноуглеродных радикалов: метила, гидроксиметилена, формила.
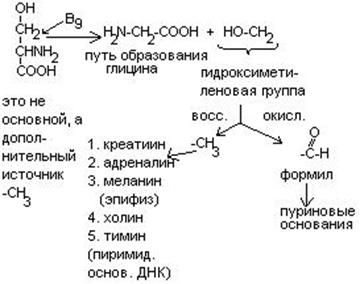
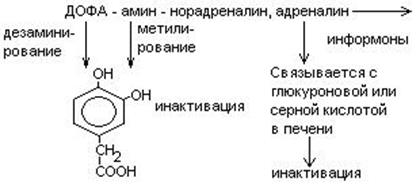
Нарушения обмена ДОФА-амина
- Паркинсонизм – мышечная дрожь, ригидность мышц (дегенерация ДОФА-амин синтезирующих нейронов). Можно моделировать паркинсонизм с помощью амфитамина
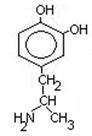
Лечат с помощью ДОФА (предшественник), легко проникает через мембраны.
- Шизофрения – избыток накопления ДОФА-амина.
Обмен цистеина и метионина.
В молекулах белка обнаружены 3 серосодержащие аминокислоты: метионин, цистеин, цистин.
Цистеин в организме синтезируется из метионина.
Функции цистеина:
- Цистеин участвует в образовании цистина:
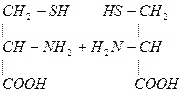
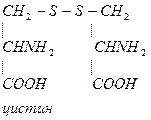
- При образовании цистина возникает дисульфидная связь S-S между двумя полипептидными цепями, что способствует стабилизации третичной структуры белка.
- Цистеин входит в состав трипептида глутатиона–Г–SH. Глутатион обеспечивает сохранение ферментов в активной форме. Глутатион участвует в ингибировании белков. Например, инсулина.
- Цистеин превращается в таурин:
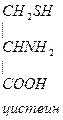
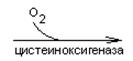
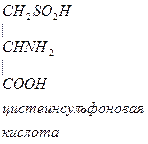
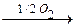
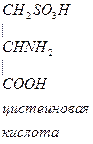
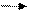
 СО2 СО2
СО2 СО2
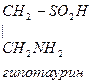
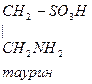
Таурин используется для синтеза парных желчных кислот.
- Цистеин входит в состав активных центров ферментов.
Метионин – незаменимая аминокислота.
Функции метионина:
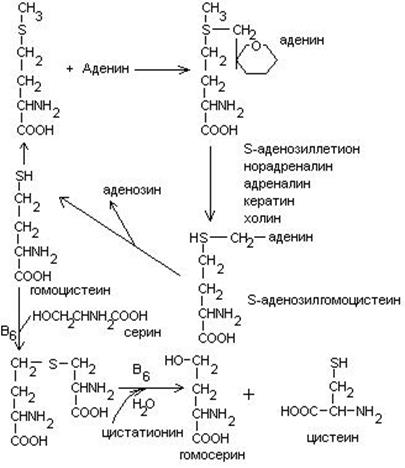
- Метионин является источником одноуглеродного радикала – метила, который используется в реакциях трансметилирования. Непосредственным источником метильных групп является производное метионина – S-аденозилметионин.
- Метионин участвует в синтезе креатина. Синтез креатина происходит в печени и почках. В почках образуется гуанидинацетат из аргинина и глицина:
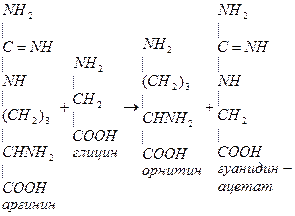
В печени гуанидинацетат взаимодействует с S-аденозилметионином и образуется креатин:
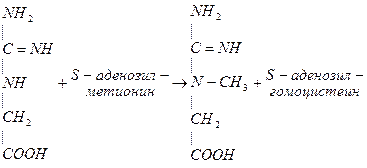
- Метионин участвует в реакциях трансметилирования в синтезе: адреналина, мелатонина, азотистых оснований.
В результате реакции трансметилирования образуется гомоцистеин.
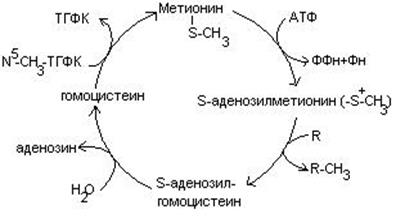
4. Гомоцистеин превращается в цистеин:
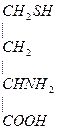 +
+ 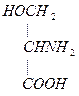
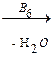
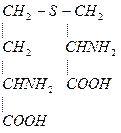
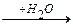
мет гомоцистеин серин цистатионин
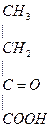 + NH3 +
+ NH3 + 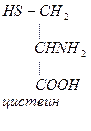
В качестве кофермента цистатионин- 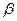 -синтаза и цистатионин-
-синтаза и цистатионин- 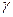 -лиаза содержат пиридоксальфосфат (В6).
-лиаза содержат пиридоксальфосфат (В6).
Гомоцистеин может превращаться в метионин путем метилирования.
Гомоцистеин может превращаться в гомоцистин. Накопление гомоцистина в тканях и крови – характерный симптом наследственной недостаточности ферментов, а также признак недостаточности витаминов В6 и В12. Гомоцистин накапливается, если нарушается превращение гомоцистеина в метионин и цистеин.
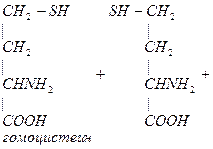
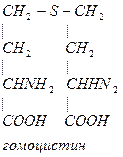
Гомоцистинурия – высокая концентрация гомоцистина и меионина. Нарушение умственного развития и скелета.
Цистинурия- выделение цистина с мочой: цистиновые камни в мочевых путях.
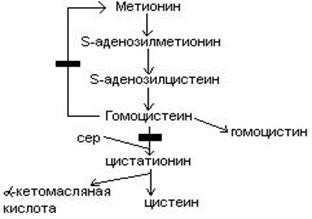
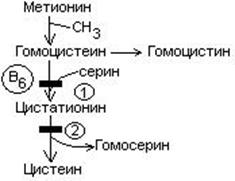
выделяют 2 причины образования блоков:
- Наследственная недостаточность ферментов, участвующих в обмене.
- Недостаточность (гиповитаминоз) В6, В12, фолиевой кислоты.
При блоке 1 развивается гомоцистинурия.
Биохимически накапливается мет и гомоцистин, т.к. гомоцистеин не превращается в цистатионин. Гомоцистин выделяется с мочой.
Клинически: нарушения со стороны соединительной ткани, сердечнососудистой системы, свертывающей системы, образование тромбов. Уродства черепа – башневидный череп, вывернутые глазные яблоки (подвывих хрусталика), умственная отсталость.
При блоке 2 развивается цистатионинурия.
Биохимически повышается концентрация цистатионина, уменьшается цистеина.
Цистинурия – в моче выделяется в 50 раз больше нормы цистина, лизина, аргинина, орнитина.
Причиной заболевания является нарушение реабсорбции цистина и нарушением всасывания остальных аминокислот.
Для людей с этой патологией характерно образование камней.
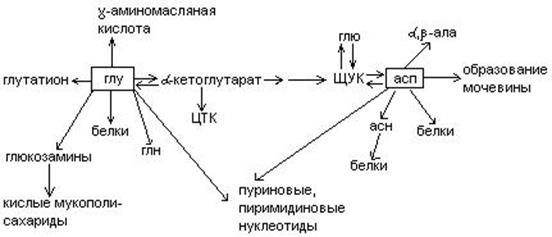
Обмен дикарбоновых аминокислот.
Глутаминовая кислота – моноаминодикарбоновая, заменимая, глюкогенная.
- Необходима для трансдезаминирования аминокислот:

- Связывает NH3 в нервных клетках, прямо на месте, по мере образования:
+ NH3 + АТФ 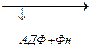
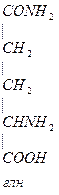
Глн входин в полипептидную цепь белка, это продукт обезвреживания NH3.
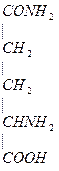
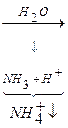
Это процесс сохранения кислотно-щелочного равновесия.
Основной щелочной эквивалент Na+, его нельзя терять с мочой. Na+ реабсорбируется, процесс регулируется гормонально. Вместо Na+ выводится 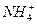 .
.
- Глутамин является возбудительным медиатором, в процессе метаболизма превращается в тормозной медиатор.
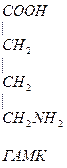
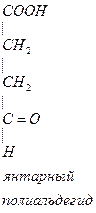
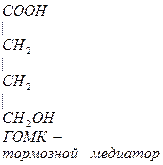
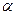 -аланин – заменимая, глюкогенная АМК, много в машечной ткани.
-аланин – заменимая, глюкогенная АМК, много в машечной ткани.
Здесь существует 2 цикла глю – лак и глю – ала.
Пируват аминируется или трансаминируется и образуется ала. Благодаря этому:
- меньше образуется лактата
- связывается определенное количество NH3 (его много в работающей мышце за счет дезаминирования).
В виде аланина NH3 идет в печень, где обезвреживается путем синтеза мочевины.
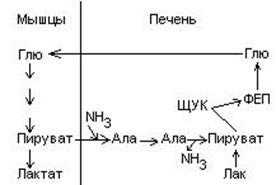
Аргинин – диаминомонокарбоновая кислота, заменимая, гликогенная.
-
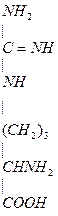 Отдача гуанидиновой группы в виде NH2-CО-NH2 (мочевины)
Отдача гуанидиновой группы в виде NH2-CО-NH2 (мочевины) - Отдача на гли гуанидиновой группы и образование креатина. При этом образуется орнитин NH2-(CH2)3-CH(NH2)-COOH. Его мало в пищевых продуктах, он не входит в белки, но он необходим для синтеза мочевины.
Основа для синтеза орнитина – аргинин.
Аспарагиновая кислота – моноаминодикарбоновая кислота, заменимая, гликогенная.
1. 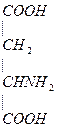 Участвует в обезвреживании NH3 с образованием аспарагина (связыв. NH3 в 10 раз меньше, чем глу). Асн входит в полипептидную цепь (в геноме есть триплет).
Участвует в обезвреживании NH3 с образованием аспарагина (связыв. NH3 в 10 раз меньше, чем глу). Асн входит в полипептидную цепь (в геноме есть триплет).
2. При дезаминировании превращается в ЩУК.
Два источника ЩУК
а) асп (энергонезависимый путь)
б) Карбоксилирование пирувата (энергозависимый путь).
Без ЩУК не идет ЦТК, невозможен для многих веществ путь превращения в глю, т.е. глюконеогенез.
3. Асп – основа для синтеза пиримидиновых оснований. 4 положения из асп кислоты (по Лениджиру только 3) 2 остальных положения из карбомаилфосфата. Обезвреживание NH3 связано с синтезом пиримидиновых оснований.
4. Асп  источник и -аланина (встречается в КоА, кранозин, ансериндипептиды – находится в мышечной ткани.)
источник и -аланина (встречается в КоА, кранозин, ансериндипептиды – находится в мышечной ткани.)
В других тканях таких дипептидов мало, нужны для функционирования мышечной ткани и повышения физической работоспособности.
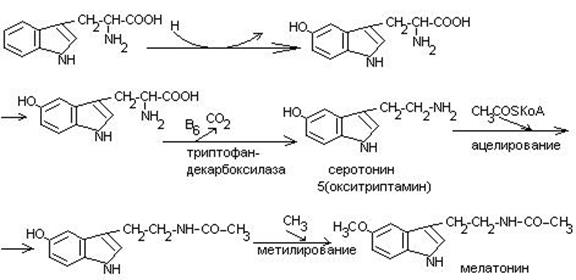
Обмен триптофана.
Триптофан – незаменимая кислота.
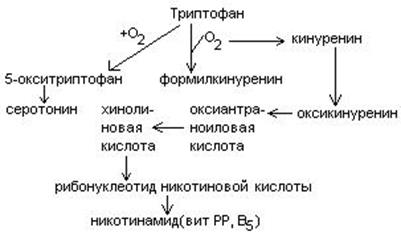
В физиологических условиях 95% триптофана окисляются по кинурениновому пути и не более 1% по серотониновому.
Основной обмен триптофана приводит к синтезу НАД, уменьшая потребность организма в витамине РР.
Обмен фенилаланина и тирозина.
Фен – незаменимая аминокислота. Тирозин может синтезироваться из фенилаланина. Глюко-кетогенные аминокислоты.
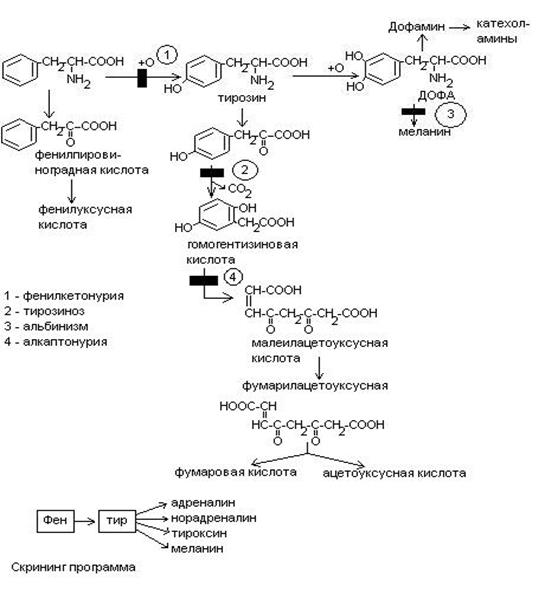
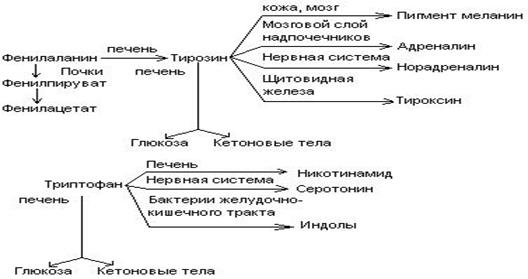
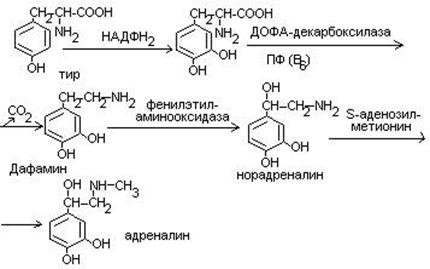
Синтез катехоламинов (адреналина, норадреналина)
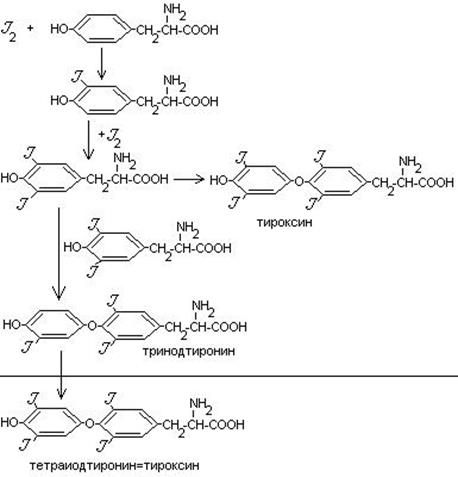
Синтез тироксина.
Обмен разветвленных аминокислот.
Вал, лей, илей.
Незаменимые аминокислоты.
Вал глю (пропионил-КоА сукцинил-КоА глю)
Лей кетокислота
Илей глю + кето (ацетил-КоА + пропионил-КоА)
фумарат
глю
Лейцин
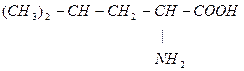

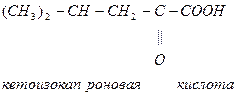
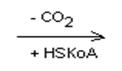
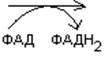
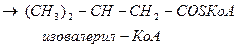


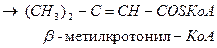
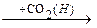

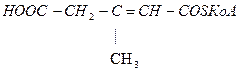
 -ОМГ-КоА
-ОМГ-КоА
Ацетоацетат Ацетил-КоА
Структура и свойства нуклеопротеидов.
Функция нуклеопротеидов заключается в хранении и передаче наследственной информации.
Состоят из белков и нуклеиновых кислот. Простетической группой нуклеопротеидов является нуклеиновая кислота.
При легком гидролизе белок дает пептиды, а нуклеиновые кислоты дают нуклеотиды или нуклеозиды.
При жестком гидролизе образуются аминокислоты, азотистые основания (аденин, гуанин, урацил, цитозин, тимин), рибоза, дезоксирибоза.
Дата добавления: 2015-08-04; просмотров: 6082;