ГЛАВА II. Классический генетический анализ
Генетический анализ - это система наблюдений и опытов, которая ставит целью вскрытие генотипической структуры особи, популяции или вида. Классическими методами генетического анализа считаются генеалогический - анализ родословных - у человека и гибридологический метод - постановка скрещиваний - у всех остальных живых существ. Основанием для проведения генетического анализа является установление факта наследования признака.
2.1 Моногенные различия
Вторым после установления факта наследования признака этапом генетического анализа является выяснение числа генов, определяющих альтернативное проявление признака. Для выяснения числа генов, вовлеченных в формирование признака необходимо определить число фенотипических классов у гибридов первого поколения и, зачастую, у потомков возвратного скрещивания (т.е. скрещивания с одной из родительских форм). Гибридами считаются потомки скрещивания особей с разным генотипом. Скрещивание родительских форм, различающихся аллелями одного гена, называется моногибридным. Различия таких форм называются моногенными. Диплоидный организм (с двойным набором хромосом), в геноме которого присутствуют два одинаковых аллеля одного гена, называется гомозиготным. Гетерозиготным является диплоидный организм с разными аллелями одного гена. Гемизиготной называют диплоидную особь, имеющую только один из аллелей данного гена.
Родительские формы обозначаются латинской буквой P (от лат. parento - родители), гибриды - буквой F (fillii - дети). Гибриды первого поколения - F1, второго - F2 и т.д. Гибриды, полученные от возвратного скрещивания - Fb (от англ. backcross - возвратное скрещивание). Если при возвратном скрещивании используется рецессивная родительская форма, такое скрещивание называется анализирующим, а его потомки обозначаются Fa (от англ. analyse - анализ). Женский пол обозначается астрологическим знаком Венеры - ♀, а мужской - знаком Марса - ♂. Скрещивание обозначают знаком умножения - X. Обычно первыми записывают особей женского пола, поскольку материнство всегда является установленным фактом, а отцовство имеет вероятностный характер. Например, P ♀ AA X ♂ Aa - скрещивание гомозиготной самки с гетерозиготным самцом. Мендель предложил записывать доминантных гомозигот двумя прописными буквами (АА), гетерозигот - одной прописной и одной строчной буквами (Аа), а гомозиготных рецессивов - двумя строчными буквами (аа). Для обозначения доминантного фенотипического класса, включающего доминантных гомозигот АА и гетерозигот Аа, используется обозначение А-. Понятно, что рецессивный фенотипический класс, включающий только гомозиготных рецессивов, обозначается аа.
Итак, по Менделю:
P ♀ AA X ♂ aa
F1 Aa
F2 3 A- : 1 aa (1 АА : 2 Аа :1 аа)
FА 1 A- : 1 aa (1 Аа : 1 аа)
Следует обратить внимание, что в случае гибридов второго поколения (F2) доминантный фенотипический класс А- представлен двумя генотипическими классами - АА и Аа - в соотношении 1 : 2, а в случае анализирующего скрещивания - только гетерозиготами Аа.
Таким образом, наличие расщепления 3 : 1 во втором поколении, и 1 : 1 в анализирующем скрещивании однозначно указывает на моногенные различия родительских форм.
Аллель, наиболее часто встречающийся в популяции, называется аллелем дикого типа, а более редкие аллели - мутантными. Аллель дикого типа обозначают буквенным символом гена со значком +, например: s+, w+.
Степень проявления признака у отдельных носителей определенного генотипа называется экспрессивность, а частота проявления признака среди носителей определенного генотипа – пенетрантность.
2.2 Типы взаимодействия аллелей
Существует пять типов взаимодействия аллелей - доминирование, неполное доминирование, кодоминирование, сверхдоминирование и межаллельная комплементация.
Классическим случаем взаимодействия аллелей, описанным Менделем, является доминирование. При этом у гибридов первого поколения проявляется признак только одной родительской формы, которая считается по этой причине доминантной (от лат. dominatio - господство). Признак родительской формы, «исчезающий» у гибридов первого поколения - рецессивный (от лат. recessus - отступление, удаление).
Пример:
При скрещивании короткошерстных (LL) и длинношерстных (ll) кошек наблюдаем следующую картину:
P ♀ LL X ♂ ll
F1 Ll (короткошерстные)
F2 3 L- : 1 ll (3 части короткошерстных и 1 часть длинношерстных)
FА 1 L- : 1 ll (1часть короткошерстных и 1 часть длинношерстных)
При неполном доминировании у гибридов первого поколения наблюдается проявление признака, промежуточное между двумя родительскими формами, а во втором поколении и у потомков анализирующего скрещивания расщепление по фенотипу полностью повторяет расщепление по генотипу.
P ♀ AA X ♂ aa
F1 Aa
F2 1 АА : 2 Аа :1 аа
FА 1 Аа : 1 аа
Пример:
При скрещивании черных (BB) и белых (bb) кур получаем в F1 голубых (андалузских) птиц. Во втором поколении - 1 часть черных, 2 части андалузских и 1 часть белых кур.
При кодоминировании два или более аллелей доминируют по отношению к рецессивной. Наиболее известный пример - наследование групп крови системы AB0 у человека.
- I группа крови соответствует генотипу I0 I0;
- II группа крови - IA IA или IA I0;
- III группа крови - IB IB или IB I0;
- IV группа крови - только IA IB.
Биохимический механизм типов взаимодействия аллелей следующий:
- при полном доминировании проявление признака определяется присутствием одного из двух доминантных аллелей, каждый из которых кодирует полнофункциональный белок, необходимый для проявления признака;
- при неполном доминировании проявление признака является дозозависимым - половина количества функционального белка приводит к частичному проявлению признака;
- при кодоминировании две или более активные формы белка, кодируемые кодоминирующими аллелями, дают возможность проявиться соответствующему признаку независимо от наличия рецессивного аллеля.
Очень редко наблюдаются явления сверхдоминирования (когда гибриды первого поколения превосходят по степени проявления признака обе родительские формы) и межаллельной комплиментации (когда у гибридов первого поколения появляется новый признак). Интересно, что биохимические механизмы сверхдоминирования и межаллельной комплементации близки – в обоих случаях аллели содержат мутации в участках, кодирующих разные домены белковых продуктов, которые являются функциональными аналогами. Объединение аллелей у гибридов первого поколения приводит к появлению продукта с новыми свойствами: большей степенью проявления признака (сверхдоминирование) или новой формой проявления признака (межаллельная комплементация).
Примеры:
- активность алкогольдегидрогеназы у гетерозиготных дрозофил выше, чем у обеих гомозиготных родительских форм;
- при скрещивании форм льна с белыми и розовыми цветками получаются растения с голубой окраской лепестков венчика.
Если присутствует серия множественных аллелей, то они последовательно проявляют по отношению друг к другу доминантность или рецессивность. Например, в локусе агути у собак присутствует следующая иерархия доминирования: ay > aw > at > a (соболиный окрас шерсти > зонарный окрас > черно-подпалый окрас > черный окрас).
2.3 Генеалогический метод
Поскольку эксперименты на людях категорически неприемлемы, генетический анализ у человека традиционно был основан на использовании генеалогического метода, который позволяет систематизировать наблюдения путем составления и изучения родословных. Как правило, родословные составляют на основе опросов, хотя по мере развития медицинской статистики и увеличения охвата населения системой медико-генетического консультирования все большую роль в этом играют компьютерные базы данных.
Генеалогическое древо - родословная, которая построена начиная от индивидуума, с которого начато исследование - пробанда, и включает всех родственников по нисходящей линии. Пробандом не обязательно является лицо, страдающее наследственным заболеванием - им может быть любой человек, обратившийся в медико-генетическую консультацию или просто участник опроса. Сибсами называются полнокровные братья и сестры, полусибсами - братья и сестры, имеющие одного общего родителя. На рисунках II, 1; II, 2 и II, 3 представлены основные обозначения, которые рекомендованы для составления родословных. При этом следует отметить, что единых правил составления родословных не существует и все обозначения должны быть отражены в легенде - списке использованных символов.

Рисунок II, 1. Обозначения индивидуумов в родословных. 1 - мужчина, 2 - женщина, 3 - пол не определен, 4 - умершая женщина, 5 - умерший мужчина, 6 - мужчина - носитель признака, 7 - женщина - носитель признака, 8 - гетерозиготный мужчина, 9 - гетерозиготная женщина, 10 - мужчина - пробанд, 11 - женщина - пробанд.
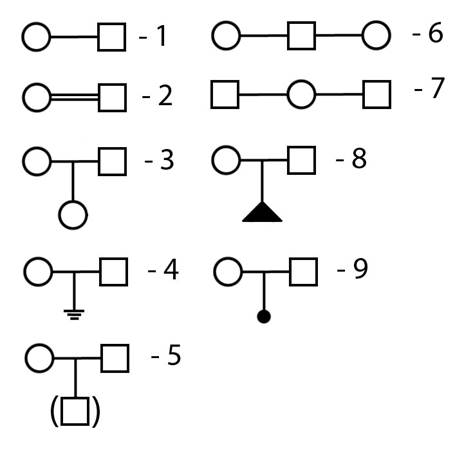
Рисунок II, 2.Обозначения семей в родословных. 1 - брак (связь), 2 - кровнородственный брак, 3 - семья с одним ребенком - девочкой, 4 - бесплодный брак, 5 - семья с усыновленным ребенком - мальчиком, 6 - связь одного мужчины с двумя женщинами, 7 - связь одной женщины с двумя мужчинами, 8 - выкидыш, 9 - медицинский аборт.

Рисунок II, 3. Обозначения близнецов в родословных. 1 - разнояйцовые (гетерозиготные) близнецы - девочки, 2 - разнояйцовые (гетерозиготные) близнецы - мальчики, 3 - разнояйцовые (гетерозиготные) близнецы - мальчик и девочка, 4 - однояйцовые (монозиготные) близнецы - девочки, 5 - однояйцовые (монозиготные) близнецы - мальчики.
2.4 Типы наследования
Аутосомно-доминантный тип наследования признаков характеризуется проявлением признака во всех поколениях (без «проскока») и у обоих полов примерно с одинаковой частотой встречаемости (Рисунок II, 4). Ген, определяющий признак, находится в одной из 22-х аутосом (то есть тех хромосом, которые одинаковы у обоих полов), доминирование полное, мутантным является доминантный аллель, аллель дикого типа - рецессивный.
Примеры:
- свободная мочка уха по отношению к приросшей мочке;
- семейная гиперхолестеринемия;
- ахондроплазия.
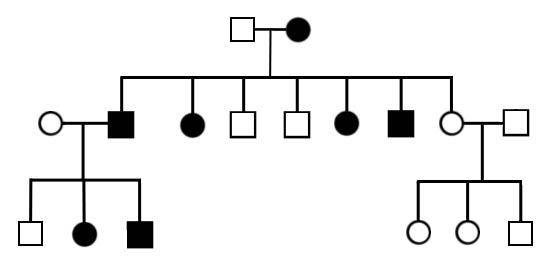
Рисунок II, 4. Аутосомно-доминантный тип наследования.
Аутосомно-рецессивный тип наследования признаков характеризуется проявлением признака у потомков родителей, которые не имели его - наблюдается «проскок поколений» (Рисунок II, 5). Представители обоих полов одинаково часто встречаются среди обладателей такого признака. Ген, определяющий признак, находится в одной из 22-х аутосом, доминирование полное, мутантным является рецессивный аллель, аллель дикого типа - доминантный.
Примеры:
- муковисцедоз;
- фенилкетонурия;
- андро-генитальный синдром.

Рисунок II, 5. Аутосомно-рецессивный тип наследования. Виден «проскок» поколений.
Сцепленное с полом доминантное наследование имеет сходство с аутосомно-доминантным - признак проявляется во всех поколениях, без «проскоков», но у женщин в два раза чаще, чем у мужчин (Рисунок II, 6). Однако в этом случае от отцов признак может передаться только дочерям, а от матерей - с равной вероятностью сыновьям и дочерям. Степень проявления признака у гетерозиготных женщин, как правило, ниже, чем у гемизиготных мужчин, что во многом объясняется инактивацией одной из X-хромосом у женщин. Ген, определяющий признак, находится в половой X-хромосоме, доминирование полное, мутантным является доминантный аллель, аллель дикого типа - рецессивный.
Примеры:
- витамин-Д-резистентный рахит с гипофосфатемией;
- рото-лице-пальцевый синдром.

Рисунок II, 6. Сцепленный с полом доминантный тип наследования.
В случае сцепленного с полом рецессивного наследования, подобно аутосомно-рецессивному, могут появляться дети - обладатели признака у не имеющих этого признака родителей и часто наблюдается «проскок» поколений (Рисунок II, 7). Однако признак передается от отца к половине дочерей, если мать гетерозиготна (проявляется только в гомозиготном состоянии), и от гетерозиготной матери к половине сыновей (проявляется всегда, так как сыновья - гемизиготы). Никогда признак не передается от отца к сыну. Ген, определяющий признак, находится в половой X-хромосоме, доминирование полное, мутантным является рецессивный аллель, аллель дикого типа - доминантный.
Примеры:
- гемофилия А;
- синдром Леша-Нихена;
- дальтонизм.
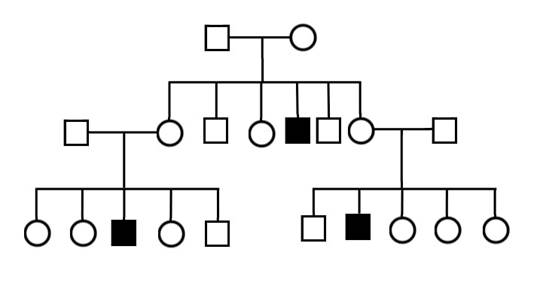
Рисунок II, 7. Сцепленный с полом рецессивный тип наследования.
Аутосомный, ограниченный полом тип наследования наблюдается в случаях аутосомной локализации гена, который его определяет, и физической возможности его проявления у особей только одного пола (процент белка в молоке или форма проявления вторичных половых признаков) (Рисунок II, 8). Этот тип наследования похож на сцепленное с полом рецессивное наследование. Главное отличие - при сцепленном с полом рецессивном наследовании признак никогда не передается от отца к сыну.
Пример:
Моно- и билатеральный крипторхизм.
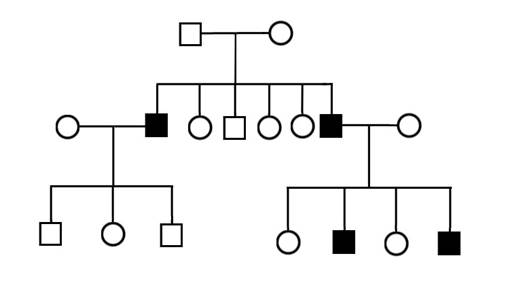
Рисунок II, 8. Аутосомный, ограниченный полом, тип наследования.
Голандрический тип наследования проявляется, если ген, определяющий признак, находится в Y-хромосоме. Поскольку в норме у мужчин только одна Y-хромосома, он всегда находится в гемизиготном состоянии. Все сыновья носителя такого признака также будут обладать им, а дочери - никогда (Рисунок II, 9).
Пример:
- волосатые уши;
- оволосение средней фаланги пальцев.

Рисунок II, 9. Голандрический тип наследования.
Митохондриальный тип объясняется цитоплазматической наследственностью, когда ген, определяющий признак находится в геноме митохондрий. Поскольку при слиянии половых клеток от сперматозоида остается только пронуклеус (гаплоидное ядро), а вся цитоплазма оплодотворенной зиготы происходит от яйцеклетки, митохондриальный тип наследования означает передачу признака от матери ко всем ее потомкам (Рисунок II, 10). Мутации митохондриального генома, как правило, приводят к тяжелым нарушениям обмена веществ.
Примеры:
- митохондриальная миоэкзенцефалия;
- атрофия зрительного нерва Лебера;
- болезнь Кернса - Сейра.

Рисунок II, 10. Митохондриальный тип наследования.
2.5 Полигенные различия
Если родительские формы различаются по двум признакам, то их скрещивание называется дигибридным. В этом случае говорят о дигенных различиях родительских форм. Если родительские формы различаются по трем и более признакам, то их скрещивание называется полигибридным. В этом случае говорят о полигенных различиях родительских форм.
P ♀ AAВВ X ♂ aaвв
F1 A-В- (по генотипу - АаВв)
F2 9 А-В- : 3 А-вв : 3 ааВ- : 1 аавв
FА 1 АаВв : 1 Аавв : 1 ааВВ : 1 аавв
Пример:
При скрещивании гороха с желтыми гладкими семенами (ААВВ) и зелеными морщинистыми (аавв) Мендель получил в первом поколении все семена - желтые гладкие (А-В-, по генотипу - дигетерозиготы АаВв), во втором поколении 9 желтых гладких (А-В-) : 3 желтых морщинистых (А-вв) : 3 зеленых гладких (ааВ-) : 1 зеленых морщинистых (аавв).
Для выяснения генотипа потомков, исходя из генотипов родителей, служит решетка Пеннета. В верхней строке выписывают гаметы одного родителя, в левом столбце - гаметы другого. Во внутренних ячейках получаем генотипы потомков.
| АВ | Ав | аВ | ав | |
| АВ | ААВВ | ААВв | АаВВ | АаВв |
| Ав | ААВв | ААвв | АаВв | Аавв |
| аВ | АаВВ | АаВв | ааВВ | ааВв |
| ав | АаВв | Аавв | ааВв | аавв |
Иногда признак определяется аллелями двух и более генов. Тогда дигенные и полигенные различия характеризуют особей, различающихся по аллелям двух и более генов. Независимое комбинирование генотипов возможно только если изучаемые гены расположены на разных хромосомах.
2.6 Взаимодействие генов
Продукты генов могут по-разному взаимодействовать. В этом случае проявление признака будет определяться аллелями двух и более генов.
Если продукты двух и более генов дополняют действие друг друга, приводя к формированию нового признака, такой тип взаимодействия генов называется комплементарность.
Пример:
У плодовой мушки Drosophila melanogaster коричневая окраска глаз определяется рецессивной мутацией bw, а аллелем дикого типа является bw+. Ярко алые глаза наблюдаются у гомозигот по мутантному аллелю st, в то время как присутствие в генотипе одного аллеля дикого типа st+ приводит к формированию нормального (красного) цвета глаз. Скрещиваем дрозофил с коричневыми и ярко алыми глазами. В первом поколении наблюдается новообразование по сравнению с обеими родительскими формами - все гибриды имеют красные глаза (генотип bw+/bw st+/st). Следует отметить, что наличие новообразования всегда говорит о взаимодействии генов. Для установления генотипов и фенотипов гибридов второго поколения рисуем решетку Пеннета:
| bw+ st+ | bw+ st | bw st+ | bw st | |
| bw+ st+ | bw+/bw+ st+/st+ красные | bw+/bw+ st+/st красные | bw+/bw st+/st+ красные | bw+/bw st+/st красные |
| bw+ st | bw+/bw+ st+/st красные | bw+/bw+ st/st коричневые | bw+/bw st+/st красные | bw+/bw st/st ярко алые |
| bw st+ | bw+/bw st+/st+ красные | bw+/bw st+/st красные | bw/bw st+/st+ коричневые | bw/bw st+/st коричневые |
| bw st | bw+/bw st+/st красные | bw+/bw st/st коричневые | bw/bw st+/st коричневые | bw/bw st/st белые |
P ♀ bw/bw st+/st+ X ♂ bw+/bw+ st/st
F1 bw+/- st+/- (по генотипу - bw+/bw st+/st)
F2 9 bw+/- st+/-: 3 bw+/bw st/st : 3 bw/bw st+/- : 1 bw/bw st/st
bw+/- st+/- - красные глаза (дикий тип)
bw+/bw st/st - ярко алые глаза
bw/bw st+/- - коричневые глаза
bw/bw st/st - белые глаза
Биохимическое объяснение состоит в том, что цвет глаз у дрозофилы определяется взаимодействием коричневого и ярко алого пигментов. Мутация bw блокирует синтез ярко алого пигмента, а мутация st - коричневого. У гомозигот по обеим мутациям bw/bw st/bw нет пигмента вообще, и, как следствие, глаза белые. У гомозигот по bw нет ярко алого пигмента - глаза коричневые (при условии наличия хотя бы одного доминантного аллеля st+), а у обладателей генотипов bw+/bw+ st/st и bw+/bw st/st - ярко алые.
При комплементарном взаимодействии генов возможны расщепления по фенотипу 9 : 3 : 3: 1, 9 : 3 :4, 9 :7.
Примерами комплементарного взаимодействия генов у человека являются врожденная глухота и наследование иммунного ответа к синтетическим полипептидам.
Другим типом взаимодействия генов является эпистаз, когда действие одного гена подавляет действие другого.
Пример:
При скрещивании белых кур и белых петухов, имеющих различное происхождение, в первом поколении все птицы были белые, а во втором поколении - 13 частей белых и 3 части окрашенных. Появление новообразования говорит о взаимодействии генов, а соотношение 13 : 3 (общее число частей - 16) является видоизменением соотношения 9 : 3 : 3 : 1, что говорит о дигенных различиях. Наличие окраски у кур определяется доминантным аллелем C, у гомозигот сс окраска белая. Доминантный ингибитор окраски I действует независимо от генотипа по гену C, подавляя формирование окрашенных перьев.
P ♀ IICC X ♂ iicc
F1 I-C- (по генотипу - IiCc)
F2 13 белые (9 I-C- + 3 I-cc +1 iicc) : 3 окрашенным (3 iiC-)
Частным случаем эпистаза является супрессия, когда подавляющим действием обладает рецессивный аллель эпистатирующего гена. При доминантном эпистазе расщепление 13 частей мутантных особей : 3 частям особей дикого типа (частный случай 12 : 3 : 1), а при супрессии - 13 частей особей дикого типа : 3 частям мутантных особей.
У человека эпистатические взаимодействия генов наблюдаются, например, при наследовании склонности к ожирению, предрасположенности к склерозу, предрасположенности к риноконьюктивиту.
Полимерным называется взаимодействие генов, которое изменяет проявление признака количественно. То есть при совместном действии двух или более генов наблюдается изменение степени выраженности признака.
Различают некумулятивную и кумулятивную полимерию. В первом случае фенотипические отличия отмечаются только у гомозиготных рецессивов по двум генам (расщепление 15 : 1), а во втором случае у гибридов второго поколения присутствуют градуальные различия (1 : 4 : 6 : 4 : 1) пропорционально числу доминантных аллелей любого из двух генов-участников. Гены, взаимодействующие по типу полимерии, обычно обозначают одинаковыми буквами с разными цифровыми индексами - A1, A2, A3.
Примеры:
- Некумулятивная полимерия наблюдается, например, при наследовании признака форма плода у пастушьей сумки (Capsella bursa-pastoris). При скрещивании родительских форм с треугольными плодами (генотип A1A1A2A2) и овальными плодами (генотип a1a1a2a2) в первом поколении гибриды имеют треугольные плоды, во втором наблюдается расщепление 15 частей растений с треугольными плодами : 1 часть растений с овальными плодами. Для формирования рецессивного фенотипа необходимо полное отсутствие доминантных аллелей обоих генов - A1 и A2.
P ♀ A1A1A2A2 X ♂ a1a1a2a2
F1 треугольные плоды (по генотипу - А1а1A2a2)
F2 15 треугольные плоды (все генотипы кроме a1a1a2a2) : 1 овальные плоды (a1a1a2a2)
- Признак окраска кожи у человека наследуется по типу кумулятивной полимерии. Наличие четырех доминантных аллелей (A1A1A2A2) приводит к формированию черной окраски кожи. В генотипе темных мулатов присутствуют три любых доминантных аллеля этой серии (то есть A1 или A2 в любой комбинации). Для формирования окраски кожи, свойственной средним мулатам, достаточно двух доминантных аллелей, а светлых мулатов - одного. Наконец, белый цвет кожи наблюдается у гомозиготных рецессивов по двум генам - генотип a1a1a2a2. На одном из островов у побережья Африки пираты провозгласили свободную республику. Все мужчины имели белую окраску кожи, а женщины – освобожденные рабы – черную:
P ♀ A1A1A2A2 X ♂ a1a1a2a2
F1 средние мулаты (по генотипу - А1а1A2a2)
F2 1 черные (A1A1A2A2) : 4 темные (2 A1A1A2а2 + 2 A1а1A2A2) : 6 средние (2 A1A1а2а2 + 2 а1а1A2A2 + 2 A1а1A2а2) : 4 светлые (2 а1а1A2а2 + 2 A1а1а2а2) : 1 белые (a1a1a2a2)
Во втором поколении только 1/8 потомков имеет фенотип одной из родительских форм и 3/8 – фенотип гибридов первого поколения. Оставшиеся 1/2 - обладатели новых фенотипов по признаку окраска кожи.
Иногда один ген имеет влияние на два и более признака. Такое явление называется плейотропией. Например, альбинизм у человека часто связан с ухудшением слуха, рыжая окраска волос – с более светлым цветом кожи и появлением веснушек, серповидноклеточная анемия – с устойчивостью к малярии. Особенно интересно рецессивное летальное (приводящее к смерти) действие некоторых доминантных мутаций (мраморный окрас у собак породы колли, укороченный хвост у кошек породы мэнкс, сложенные уши у кошек породы скоттиш фолд). В этом случае признак проявляется только у гетерозигот, а мутантный аллель в гомозиготном состоянии приводит к гибели.
2.7 Сцепленное наследование
Два типа наследования - сцепленное с полом доминантное и сцепленное с полом рецессивное - объясняются локализацией исследуемых генов в половой X-хромосоме. Иными словами - пол, рассматриваемый как простой признак, и другой изучаемый признак наследуются совместно. Сцепление с полом - частный случай сцепленного наследования, смысл которого заключается в нарушении принципа независимого наследования двух и более признаков по причине нахождения обуславливающих их генов в одной хромосоме.
На вопрос о том, является ли наследование сцепленным с полом, могут ответить реципрокные скрещивания - когда родительские формы меняют местами. Например, вначале доминантной родительской формой являются самки, затем самцы. Различные результаты реципрокных скрещиваний говорят о сцеплении признака с полом.
Пример:
Белая окраска глаз у дрозофилы может определяться мутантной аллелем w. Ген w локализован в X-хромосоме и может быть представлен аллелем дикого типа w+ (красная окраска глаз) и мутантным аллелем w. Гомо- и гетерозиготными могут быть только самки, а самцы всегда гемизиготны по локусу w. Результаты реципрокных скрещиваний представлены на рисунке II, 11. В случае скрещивания белоглазых самок с красноглазыми самцами в первом поколении все самки красноглазые, а все самцы - белоглазые. Если скрестить красноглазых самок с белоглазыми самцами, то все потомки независимо от пола будут иметь красную окраску глаз.

Рисунок II, 11.Реципрокные скрещивания для установления типа наследования белой окраски глаз у Drosophila melanogaster.
Если два или более признаков определяются генами, локализованными на одной аутосоме, то говорят об аутосомном сцеплении (или просто сцеплении). Сцепление может быть полным - когда родительские комбинации аллелей всегда передаются потомкам, что объясняется отсутствием обмена участками хромосом (кроссинговера) в районе между исследуемыми генами. Предположим, между генами А и В - полное сцепление, тогда:
P ♀ AAВВ X ♂ aaвв
F1 АаВв
Гибриды первого поколения образую следующие гаметы:
| АВ | ав | |
| АВ | ААВВ | АаВв |
| ав | АаВв | аавв |
F2 1 ААВВ : 2 АаВв: 1 аавв
Если на участке между А и В происходит кроссинговер (рекомбинация), то кроме родительских сочетаний аллелей, например, АВ и ав будут появляться и рекомбинантные сочетания: Ав и аВ. Гаметы, несущие рекомбинантные сочетания аллелей называются кроссоверными гаметами, а происшедшие в результате их слияния особи - кроссоверными особями. Соотношения кроссоверных и некроссоверных особей зависит от расстояния на хромосоме между изучаемыми генами - чем дальше они друг от друга, тем чаще случается кроссинговер на участке между местами их локализации и тем больше кроссоверных особей. Рекомбинационное расстояние между двумя генами – это отношение числа кроссоверных гамет к общему числу гамет, умноженное на сто. Единицей рекомбинационного расстояния является 1 сантиморган (сМ). Следует отметить, что в анализирующем скрещивании FА соотношение кроссоверных и некроссоверных гамет будет равно соотношению кроссоверных и некроссоверных особей. Если одна родительская форма несет два доминантных аллеля (АВ), а другая два рецессивных (ав) - это состояние притяжения, а если родительские формы несут по одному доминантному и одному рецессивному аллелю (Ав и аВ) - это состояние отталкивания.
Пример:
У дрозофилы черная окраска тела определяется аллелем b, а b+ - аллель дикого типа. Мутация pr в гомозиготе приводит к пурпурной окраске глаз, нормальный - красный цвет глаз определяется аллелем pr+. Скрещиваем черных мух с пурпурными глазами (bbprpr) и серых мух с красными глазами (b+b+pr+pr+). Обращаем внимание на то, что аллели находятся в состоянии притяжения. Все гибриды первого поколения гетерозиготны и имеют фенотип второй родительской формы. В анализирующем скрещивании получаем:
серое тело, красные глаза - 1000
серое тело, пурпурные глаза - 64
черное тело, красные глаза - 62
черное тело, пурпурные глаза - 831
Следует отметить, гомозиготных рецессивов всегда несколько меньше, чем доминантных гомозигот из-за некоторого снижения жизнеспособности.
Нетрудно рассчитать рекомбинационное расстояние между генами b и pr.
(64 + 62) : (1000 +64 + 62 +831) x 100 = 6,4 сМ
Используя большое число генов – маркеров генетического анализа – можно построить генетические карты хромосом с указанием взаимного расположения и расстояния между маркерами. Каждая хромосома будет соответствовать одной группе сцепления. Если расстояние между двумя маркерами больше 50 сМ (что соответствует проценту кроссинговера при независимом наследовании) маркеры наследуются не проявляя сцепления друг с другом. Поэтому для повышения точности генетических карт необходимо использовать как можно большее число маркеров. На точность определения генетических расстояний влияет и явление генетической интерференции – подавления кроссинговера на участках, находящихся вблизи от участка, где уже происходит рекомбинация.
Контрольные вопросы и задания к главе II
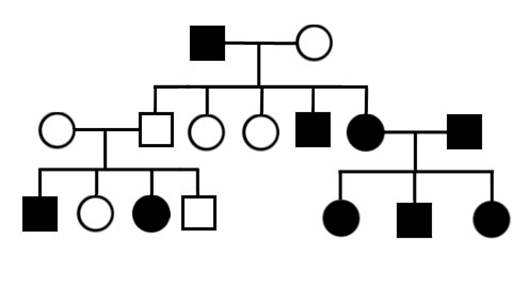
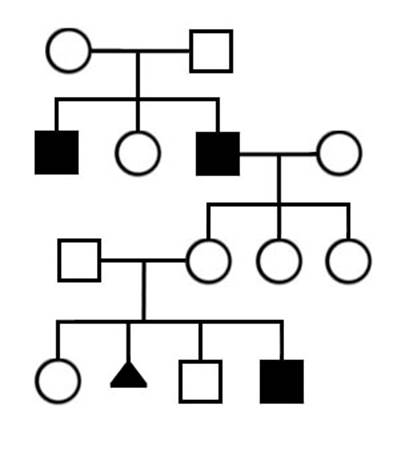
1. Определите типы наследования в родословных, приведенных на рисунке II, 12.
2.При скрещивании кошек с разной окраской шерсти было получено:
P самки черные х самцы рыжие самки рыжие х самцы черные
F1 самки черепаховые самки черепаховые
самцы черные самцы рыжие
F2 самки черепаховые самки черепаховые и рыжие
и черные самцы черные и рыжие
самцы черные и
рыжие
Определите генотипы скрещиваемых форм и локализацию генов.
3. Какие группы крови возможны у ребенка, если его родители имеют группы крови А и В, и оба являются: а) гетерозиготами, б) один из родителей гетерозиготен.
4. Потемнение зубов определяется двумя доминантными генами, один из которых находится в Х-хромосоме, а другой – в аутосоме. В семье, где родители имели темные зубы, родились девочка и мальчик с нормальным цветом зубов. Темные зубы матери определены геном, сцепленным с Х – хромосомой, а темные зубы отца – аутосомным геном. Определите генотипы родителей и детей.
5. Заболевание обнаруживается у детей, родители которых являлись двоюродными братом и сестрой и не страдали от этого заболевания. Как наследуется болезнь?
6. Известно, что ген гемофилии и ген дальтонизма – рецессивные, локализованные в X- хромосоме; расстояние между ними - 9,8 сМ. Здоровая девушка, мать которой дальтоник, а отец – гемофилик, выходит замуж за здорового мужчину, родители которого здоровы. Определить, какова вероятность появления в этой семье здоровых детей.
7. Резус–положительность и эллиптоцитоз определяются доминантными аутосомными генами. Локус резус – фактора (Д) и локус эллиптоцитоза (Е), вызывающего овальную форму эритроцитов, находятся сцепленно в одной аутосоме на расстоянии 3 сМ. Мать гетерозиготна по обоим анализируемым признакам. Отец резус – отрицателен и имеет нормальные эритроциты. Определить процентное соотношение вероятных генотипов и фенотипов детей в семье.
8. Гипертрихоз передается через Y- хромосому, а полидактилия – как аутосомный признак. В семье, где отец имел гипертрихоз, а мать – полидактилию, родилась нормальная в отношении обоих признаков дочь. Какова вероятность рождения сына без обеих аномалий?
9. У супругов с нормальным зрением родилось два сына и две дочери. У первой дочери зрение нормальное; у нее 3 сына, 2 из которых дальтоники. У второй дочери и у ее пяти сыновей зрение нормальное. Первый сын дальтоник; у него две дочери и два сына, и все видят нормально. Второй сын и четверо его сыновей также имеют нормальное зрение. Каковы генотипы всех родственников?
10. На рисунке II, 13 приведены генотипические данные особей второго поколения скрещивания дрозофил. Установите генотипы родителей.

А.
Б.
В.

Г.

Д.

Е.
Ж.
Рисунок II, 12. Примеры родословных для задачи 1 к главе II.
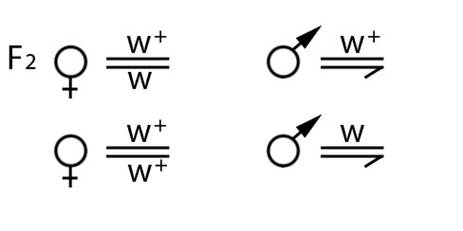
Рисунок II, 13.Генотипические данные особей второго поколения скрещивания дрозофил для задачи 10 к главе II.
Дополнительная литература к главеII
М.М. Тихомирова. Генетический анализ: учебное пособие // Л. Издательство ЛГУ. 1990. 280 С.
Дата добавления: 2015-07-24; просмотров: 2812;