Упаковка генетического материала
Если бы всю ДНК одной клетки в форме простой двойной спирали вытянуть в одну линию, то ее длина была бы 1,74 м. Представленной в виде линии суммарной ДНК одного человека можно три раза обернуть земной шар по экватору. Сильно конденсированные структуры ДНК, содержащиеся в ядрах клеток,называютсяхромосомы. При конденсации происходит уменьшение продольных размеров молекулы ДНК в десятки тысяч раз за счет образования сверхспиралей ДНК.
У эукариотических организмов значительная часть ДНК окружена множеством различных белков. Эти белки вместе с ДНК образуют комплексную структуру – хроматин, который обеспечивает специфический для эукариот тип регуляции экспрессии генов. В состав хроматина входят очень длинные двухцепочечные молекулы ДНК, белки гистоны, кислые белки и небольшие количества РНК. Всего известно пять типов гистонов: H1, H2A, H2B, H3, H4. Гистоны объединяют несколько групп основных белков. Н1 наиболее слабо связан с хроматином.
В хромосоме ДНК с помощью гистонов упакована в специальные регулярно повторяющиеся структуры – нуклеосомы. Так, образуется структура, похожая на бусы, где каждая бусина – нуклеосома. Нуклеосома представляет собой сегмент ДНК длиной около 200 пар оснований, навитый на белковую сердцевину, состоящую из восьми молекул–гистонов. В нуклеосомную сердцевину (нуклеосомный кор) входит по две молекулы гистонов H2A, H2B, H3, H4. Поверхности этих белковых молекул несут положительные заряды и образуют стабилизирующий остов, вокруг которого может закручиваться отрицательно заряженная молекула ДНК. Гистон Н1 размещается на участках ДНК, соединяющих одну нуклеосому с другой. ДНК этих участков называют соединительной (линкерной). Предполагают, что Н1 участвует в регуляции транскрипционной активности хроматина и не участвует в стабилизации структуры хромосомы.
Перед началом деления клеточного ядра хромосома, представленная на этот момент цепочкой нуклеосом (фибриллой), начинает спирализоваться, образуя хроматиновые петли и упаковываться, образуя при помощи белка H1 в толстую хроматиновую нить, или хроматиду, диаметром 30 нм. В результате дальнейшей спирализации диаметр хроматиды достигает ко времени метафазы 700 нм. Значительная толщина хромосомы (диаметр 1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой микроскоп.
Т.о. конденсированная хромосома имеет вид буквы X (часто с неравными плечами) и представляют собой высокую степень конденсации хроматина, постоянно присутствующего в клеточном ядре (рис.12).
Митотическая хромосома состоит из двух сестринских хроматид и центромеры. В зависимости от расположения центромеры хромосомы морфологически разделяют на
- Метацентрические– центромера расположена посередине хромосомы (равноплечие) – рис. 13, 1;
- Субметацентрические –центромера смещена к одному из краев (слабо неравноплечие) – рис. 13, 2;
- Акроцентрические– центромера сильно смещена к одному из плеч (резко неравноплечие) – рис. 13, 3;
- Телоцентрические– центромера расположена в теломерном (концевом) участке хромосомы – рис. 13, 4.
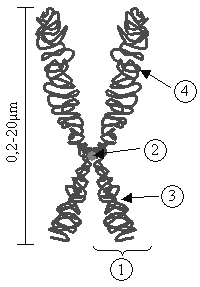
Рис. 12. Схема строения хромосомы
1—хроматида; 2—центромера; 3—короткое плечо; 4—длинное плечо.
Кроме первичной перетяжки хромосома может иметь вторичную перетяжку (Рис. 13, 5) не имеющую отношения к прикреплению нитей веретена. Местонахождение этой перетяжки в хромосоме связано с формированием ядрышка. Этот участок хромосомы называется ядрышковым (нуклеолярным) организатором. Полагают, что он имеет сложную структуру и отвечает за синтез рибосомной РНК. Иногда на концах хромосом находятся небольшие тельца – спутники. Такие хромосомы называют спутничными (рис.13, 6 ).
|
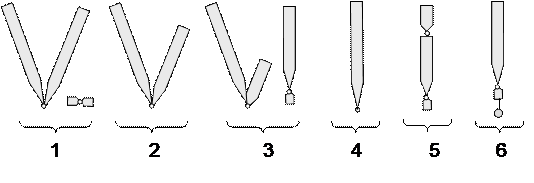
Рис.13. Схематическое строение морфологически различных хромосом
Каждая хромосома обязательно имеет центромеру, выполняющую функцию механического центра хромосомы. Именно к ней в метафазе прикрепляется нить веретена, разводящая хромосомы к полюсам. Местоположение центромеры в разных хромосомах может быть различно, оно типично для каждой хромосомы. Центромеры определяют ориентацию хромосом в митозе и их правильное расхождение к полюсам. В случае разлома хромосомы, бесцентромерный участок (ацентрический фрагмент) не может восстановить центромеру и при клеточном делении чаще всего утрачивается. Фрагмент может сохраниться лишь в том случае, если прикрепиться к хромосоме, имеющей центромеру. Центромера содержит ДНК и также является самовоспроизводящейся структурной частью хромосомы.
На концах хромосом имеются сегменты, препятствующие склеиванию хромосом концами. Такие сегменты названы теломерами. Хромосомы по своей оси неоднородны как по химическим и физическим свойствам, так и генетически. При фиксации и окраске основными красителями разные участки (районы) и даже некоторые целые хромосомы дают разную реакцию. Одни участки интенсивно окрашиваются – их называют гетерохроматиновыми, другие окрашиваются слабо. Их называют эухроматиновыми. Гетеро- и эухроматиновые участки обладают разными генетическими свойствами. Первые почти не содержат генов и наследственно инертны (соответствуют интронным областям ДНК), тогда как вторые содержат гены и наследственно активны (соответствуют экзонным областям ДНК). Гетерохроматиновые участки разбросаны по всей длине хромосомы, но чаще располагаются вблизи центромеры. Разрывы хромосом происходят как правило именно в этих участках, или на границе гетерохроматинового участка с эухроматиновым. Существуют некоторые виды хромосом, которые преимущественно содержат гетерохроматин, например, Y–хромосома.
Половыми хромосомами называют хромосомы, различно устроенные у мужских и женских организмов. По традиции, половые хромосомы, в отличие от аутосом (не половых хромосом), обозначаются не порядковыми номерами, а буквами X, Y, причём отсутствие хромосомы обозначается цифрой 0 (ноль). Как правило, при этом один из полов определяется наличием пары одинаковых половых хромосом (гомогаметный пол), а другой — комбинацией двух непарных хромосом или наличием только одной половой хромосомы. У человека, гомогаметный пол — женский (XX), гетерогаметный пол — мужской (XY).
Дата добавления: 2015-07-22; просмотров: 1769;