Морфология элементарного ландшафта 1 страница
Морфологические особенности элементарных ландшафтов изучены слабо. В некоторых работах приводятся сведения о площади и форме, вертикальном строении и мощности сделана попытка объяснить причину окраски элементарного ландшафта.
Площадь и форма элементарного ландшафта чаще всего зависят от рельефа, реже площадь — от породы. Элементарный ландшафт может занимать от нескольких квадратных метров до квадратных километров и более (низинные болота Полесья или Западной Сибири; солончаки и такыры Средней Азии). Элементарные ландшафты могут иметь круглую, эллипсовидную, полосчатую, зигзагообразную, дугообразную и некоторые другие формы. Вертикальный профиль элементарного ландшафта делится на ярусы, или горизонты (рис. 2).
В надземной части вертикального профиля элеменарного ландшафта выделен ярус живого вещества. Для этого яруса характерна концентрация элементов-органогенов (С, Н, О, N). Их соединения в летучих формах обусловливают специфический состав приземного воздуха. Мощность яруса живого вещества максимальная во влажных тропических лесах, минимальная — в водорослевых и лишайниковых сообществах пустынь.
Ниже располагается ярус почв, мощность которого определяется степенью интенсивности почвообразовательного процесса. В пределах этого яруса выделяются почвенные горизонты, которые отличаются содержанием элементов и их сочетанием. В почвенной толще протекают процессы взаимодействия между живыми организмами и органо-минеральными соединениями.
Ниже яруса почв расположена порода, где происходят физико-химические процессы, которые носят общее название — процессы выветривания. Этот ярус называется ярусом коры выветривания.
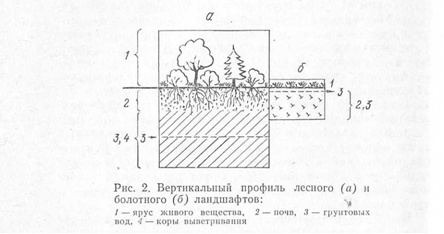
Самый нижний ярус вертикального профиля ландшафта — ярус грунтовых вод. Геохимические особенности процессов, протекающих в этом ярусе, определяются режимом и составом вод и водовмещающих пород, а также составом химических элементов, поступающих из ярусов коры выветривания и почв.
В элементарных ландшафтах могут отсутствовать отдельные ярусы. Например, иногда верхний водоносный горизонт находится за пределами ландшафта (горный элювиальный ландшафт), в других случаях он совмещен с корой выветривания или почвой (торфяно-болотная почва), в третьих — кора выветривания совмещена с почвой (горные почвы).
Окраска ландшафта — важный морфологический признак. По современным представлениям окраска — физическое явление селективного поглощения или отражения кристаллом определенного диапазона электромагнитного излучения в пределах видимой области спектра (А. Н. Платонов, 1976).
Окраски минеральных соединений по их происхождению А. Е. Ферсман (1936) впервые разделил на идиохроматические, т. е. собственные; аллохроматические, вызванные наличием в кристалле минерала второстепенных элементов — хромофоров (титан, железо, марганец, хром и др.); псевдохроматические, связанные с интерференционными явлениями.
Общая окраска ландшафта зависит от наиболее распространенных элементов и минералов, а также от органических соединений. Совокупность ионов кремния придает кварцевому песку прозрачность. Белый цвет почвенных новообразований степных и пустынных ландшафтов объясняется наличием кальция, натрия (корки солончаков, известковые конкреции). В ярусе почв и коры выветривания преобладают оттенки красного и желтого цвета как результат присутствия трехвалентного железа и гидрофильных его соединений. Зелено-голубой цвет глеевых горизонтов заболоченных почв определяется двухвалентным железом. Оттенки окраски изменяются также в зависимости от степени увлажненности породы. Влажная порода имеет более яркий и темный оттенок по сравнению с аналогичной сухой породой.
Окраску высших растений (от оранжевой до синей) создают антоцианы, которые содержатся почти во всех растительных тканях. К группе антоцианов относятся полифенолы. Цвет антоцианов определяется их структурой и физическими свойствами растворителя. С увеличением количества фенольных гидроксильных групп (ОН) цвет изменяется от розового к синему, а метилирование (СН3) гидроксильных групп — в обратном направлении. Кроме того, антоцианы меняют цвет под влиянием реакции среды, а также ионов металлов. Например, цианидин в кислом растворе имеет красный цвет, с увеличением рН интенсивность красного цвета уменьшается и при щелочной реакции цвет становится синим, а затем зеленым. После прибавления предварительно растворенного хлорида алюминия к раствору цианидина и его производных цвет раствора переходит из красного в синий.
Понижение температуры воздуха влияет на изменение цвета листьев высших растений, что объясняется накоплением в них прежде всего производных цианидина (С. С. Танчев, 1980).
Геохимический ландшафт
Обмен химическими элементами происходит в ряду основных элементарных ландшафтов: элювиальный супераквальный>субаквальный. Они составляют геохимический ландшафт (по К. Г. Раману — ряд фаций). Геохимический ландшафт может не включать некоторые подчиненные элементарные ландшафты, например субаквальный, транссупераквальный, трансэлювиальный. Ярусы коры выветривания и почв могут слагать породы монолитные (однородные по составу и генезису) и гетеролитные (разнородные). Закономерности миграции химических элементов следует изучать по геохимическому ландшафту на монолитной породе.
Если в подчиненные элементарные ландшафты поступают элементы только своего ряда фаций, то такие ландшафты называются автохтонными (геохимически подчиненными). Привнесение в подчиненный элементарный ландшафт химических элементов из других рядов определяет его как аллохтонный (геохимически слабо подчиненный).
Последовательное чередование геохимических ландшафтов (рядов фаций), сходных по условиям миграции, образует местный ландшафт, или местность. Разнообразие литологического состава пород, сочетаний почв, форм рельефа, растительных сообществ усложняет принципы классификации местных ландшафтов. М. А. Глазовской выделены простые и сложные, одноступенчатые и многоступенчатые местные ландшафты.
Простой местный ландшафт представляет собой чередование геохимических ландшафтов, идентичных по условиям миграции химических элементов, строению рельефа, генезису и составу пород и почв, истории формирования (например, плоские моренные супесчаные холмы, сменяющиеся моренными супесчаными ложбинами). Однако абсолютно одинаковые формы рельефа практически отсутствуют. Если в пределах местного ландшафта сочетаются положительные и отрицательные формы рельефа с различными морфометрическими показателями, породы и почвы пестры по составу, генезису и возрасту, а растительные сообщества разнообразны, то при едином историческом развитии такой ландшафт называется сложным местным ландшафтом. При условии существенного различия отдельных частей местного ландшафта с точки зрения их исторического развития его выделяют как сочетание местных ландшафтов.
В северной части Белоруссии встречаются моренные холмы, различные по площади и относительной высоте. В понижениях в одном случае формируется верховое болото, в другом — озеро, в третьем — разнотравно-злаковый луг, что обусловлено различными стадиями единой эволюции растительности. Породы, слагающие этот участок, однородны по составу и генезису. Все это позволяет отнести данную территорию к сложному местному ландшафту.
Для горных систем характерны многоступенчатые * простые и сложные местные ландшафты, что обусловлено особенностями рельефа. Границы многоступенчатого простого местного ландшафта выделяются по следующим признакам: элювиальные ландшафты низшей ступени геохимически независимы от вышележащих геохимических ландшафтов; супераквальные ландшафты подчинены геохимическим ландшафтам вышележащих ступеней. Если супераквальные ландшафты нижележащих ступеней рельефа не связаны миграцией элементов с элювиальными или супераквальными ландшафтами более высоких ступеней, их выделяют в сложный многоступенчатый местный ландшафт или сочетание многоступенчатых местных ландшафтов в зависимости от истории развития.
3 Широколиственные леса, влажные субтропические, тропические и экваториальные леса
Лесные ландшафты формируются в условиях избыточного увлажнения с промывным водным режимом, т. е. представлены в границах от умеренного до экваториального пояса. В этих ландшафтах хорошо выражена водная миграция элементов, которой противостоит биологический геохимический барьер. В структуре биомассы выделяется многолетняя надземная часть (более 70%), биомасса самая высокая по сравнению с другими группами ландшафтов, зольность на единицу веса растений низкая. Самой высокой продуктивностью характеризуются лесные ландшафты экваториального, субэкваториального и тропического поясов.
Экваториальные, субэкваториальные и тропические леса
* В дальнейшем лесные ландшафты экваториального, субэкваториального и тропического поясов будем называть лесными ландшафтами тропиков.
Территория, расположенная между северным и южным тропиком, характеризуется отсутствием холодного сезона. Постоянная положительная температура воздуха и различные условия увлажнения обусловливают разнообразие растительных сообществ (леса, саванны, пустыни). Лесные ландшафты экваториального, субэкваториального и тропического поясов представляют собой самостоятельные типы ландшафтов, но характеризуются сходными геохимическими условиями.
Гидротермические условия. Среднемесячная температура в течение года изменяется в экваториальном и субэкваториальном лесу от 24 до 28, в тропическом — от 18 до 28° С. Во влажных экваториальных лесах выпадает 1500—10 000 мм осадков в год, в тропических лесах— 1000—2500 мм. Испарение составляет 1000—1250, испаряемость— 1250—1500 мм в год. За исключением районов с сезонно-влажными лесами, осадки в течение года распределяются равномерно и выпадают, регулярно. Такие гидротермические условия ускоряют геохимические процессы, направленные на освобождение химических элементов из породы и миграцию их за пределы ландшафта. Геохимическим барьером являются корневая система растений, суглинистые и глинистые породы. Между элементарными ландшафтами преобладает совершенное сопряжение. Господствует промывной водный режим, который хорошо выражен на рыхлых породах. На плотных породах происходит преимущественно поверхностный перенос элементов в различных формах, хотя частично этому препятствует растительность.
Кора выветривания. Основной источник водных мигрантов в ландшафте — кора выветривания. В лесных ландшафтах жаркого пояса кора выветривания представлена магматическими, метаморфическими и осадочными породами и достигает мощности нескольких десятков и даже сотен метров. Поверхностная часть ее служит основой формирования почвенного покрова. Основные горные породы в процессе гипергенеза дают глинистые продукты, а кислые, содержащие много кварца,— песок. Основу коры выветривания ландшафтов жаркого пояса составляют устойчивые оксиды железа и алюминия, которые определяют тип химизма коры выветривания: ферритная (Fe), ферраллитная (Fe, А1), аллитная (А1). В соответствии с типом химизма коры выветривания в некоторых источниках называются почвы (ферраллитная, ферритная, аллитная).
В результате активно протекающих геохимических процессов (растворения, гидратации, окисления, восстановления, метасоматоза, изоморфизма, сорбции, парагенезиса и др.) из неустойчивых соединений, представленных преимущественно первичными минералами, образуются более устойчивые вторичные или видоизмененные минералы. Среди глинистых (вторичных) минералов наиболее устойчивым к геохимическим условиям явлется каолинит (составляет 70% всех глинистых минералов). При нейтральной реакции, которая характерна для карбонатных пород, устойчив монтмориллонит. Каолинит имеет низкую емкость поглощения ионов (1—24 мг-экв. на 100 г породы), поэтому освобождающиеся и мигрирующие элементы слабо сорбируются им и выносятся водными потоками. Тонкодисперсная часть коры выветривания представлена отчасти метагаллуазитом, гидрослюдой, смешанно-слойными минералами, аллофаноида-ми переменного состава, оксидами железа. Емкость поглощения ионов галлуазитом выше (40—60 мг-экв. на 100 г породы), чем каолинитом.
Оксид железа, сорбированный глинистыми минералами, исключая монтмориллонит, образует пленку, которая придает им красный цвет. Процесс закрепления оксида железа на поверхности каолинитовых коллоидных частиц И. А. Денисов (1971) называет хроматизацией (от греч. хрома — цвет). В горах при понижении температуры процесс хроматизации замедляется. При избытке железа образуются пленки свободного гидроксида железа, которые устойчивы к механическому воздействию. Они цементируют породу в латеритную плиту мощностью до 4—5 м и более. В латеритных панцирях могут аккумулироваться гидроксиды алюминия, марганца,  редкие и рассеянные элементы —
редкие и рассеянные элементы — 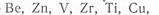
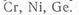 В.В. Добровольский (1975) относит латеритные панцири к гидроморфным аккумуляциям железа участков сезонного заболачивания, а зарубежные ученые — к остаточным образованиям в элювиальных ландшафтах. Изменение интенсивности окраски коры выветривания зависит от степени гидратации: чем больше гидратированы оксиды железа, тем цвет их ближе к желтому, в противном случае — к красному, бурому, красно-бурому. По сравнению с исходной породой содержание оксида железа в коре выветривания увеличивается с 6—20 до 30%.
В.В. Добровольский (1975) относит латеритные панцири к гидроморфным аккумуляциям железа участков сезонного заболачивания, а зарубежные ученые — к остаточным образованиям в элювиальных ландшафтах. Изменение интенсивности окраски коры выветривания зависит от степени гидратации: чем больше гидратированы оксиды железа, тем цвет их ближе к желтому, в противном случае — к красному, бурому, красно-бурому. По сравнению с исходной породой содержание оксида железа в коре выветривания увеличивается с 6—20 до 30%.
Алюминий в коре выветривания ландшафтов жаркого пояса образует гидраты, которые выпадают из раствора с образованием геля. Гель постепенно теряет воду и образует гиббсит (А1203 • ЗН20), бемит (А1203-Н20), диаспор (А10-ОН). В начальных стадиях выветривания породы преобладает гиббсит, который является остаточным образованием и характерен для древних латеритов. Алюминий с кремнием кристаллизуется чаще всего в форме каолинита, на породах с кальцием — в форме монтмориллонита. Содержание оксида алюминия в коре выветривания по сравнению с исходной породой увеличивается с 15—20 до 5.0%. Свободный кремнезем встречается в виде золя, а при дегидратации и кристаллизации образует вторичный кварц, или халцедон. Если аморфная структура сохраняется, то формируется опал. Кремний находится в основном в связанной форме. Растворимость соединений кремния увеличивается при рН больше 10. С ионами алюминия кремний образует распространенные глинистые минералы.
Среди редких и рассеянных элементов в коре выветривания и почвах аккумулируются титан, галлий, цирконий, которые образуют устойчивые минералы. Из коры выветривания сравнительно быстро выносятся Na, CI, S, Са, Mg, К, Mn, Sr, Ва, Li и др.
Почвы. На почвенной карте мира, составленной группой советских почвоведов, в пределах лесных ландшафтов жаркого пояса выделены следующие типы почв: а) красно-желтые (ферраллитные и альферритные) постоянно влажных лесов; б) красные (ферраллитные и альферритные) сезонновлажных лесов; в) желтые (аллитные) постоянно влажных лесов.
По-разному складывается миграция основных химических элементов в сочетании с гумусовыми кислотами. Гуминовые кислоты, связанные с кальцием и другими двухвалентными металлами, на поверхности первичных минералов образуют пленки и задерживают их разложение. Однако в почвах лесных ландшафтов тропиков господствуют фульвокислоты. В соединениях с полуторными окислами (Fe2О3 +А1203) они образуют хелаты, растворимость которых определяется величиной рН и концентрацией раствора. Например, в кислой среде фульваты железа и алюминия растворимы при равных соотношениях, а при соотношениях полуторных окислов и фульвокислот 6:1—нерастворимы. Последнее соотношение более характерно для почв тропиков. Содержание гумуса в почвах невысокое 1—4%. В анаэробных условиях происходит накопление торфа (преимущественно низинного типа, различной мощности), который удерживает химические элементы от выноса.
В почвах лесных ландшафтов поведение элементов определяется реакцией среды (рН<5), составом гумуса (преобладают фульвокислоты), окислительно-восстановительными условиями, промывным водным режимом, водно-физическими свойствами почвы, зольностью опада, емкостью поглощения почв, зарядом коллоидной частицы. Результатом этой сложной взаимосвязи является вынос за пределы почвенного профиля щелочных и щелочноземельных оснований и значительной части кремния. В почвах лесных ландшафтов тропиков преобладают положительно заряженные частицы коллоидов, поэтому характерна аккумуляция анионогенных элементов S, Р, N, Se, Ti, V и др. Мало сорбируется катионов, так как ацидоиды представлены преимущественно гумусовыми соединениями, которых в почве меньше, чем базои-дов (коллоидные частицы гидроксида алюминия и железа).
В минеральной части почвы, как и в коре выветривания, господствуют соединения алюминия и железа. Отношение Si02 : R2O3 и Si02 : А1203 меньше 2, а в иле меньше 2,5, что указывает на то, что минеральная часть почвы по составу является ферраллитной. При молекулярных соотношениях Si02 : А1203 меньше 1 почвы относятся к аллитным, приобретают цвет, близкий к желтому. Дифференциация почв на горизонты здесь слабо выражена.
Таким образом, перераспределение химических элементов в системе кора выветривания — почва носит сложный характер. Наблюдается общая тенденция к уменьшению содержания химических элементов в этих составных частях.
Воды. В ландшафтах лесов тропиков почвенно грунтовые, а также речные и озерные воды имеют, как правило, низкую минерализацию и относятся к ультрапреспым, а по химическому составу — к гидрокарбонатным и гидрокарбонатно-кремниевым. Минерализация воды в озерах резко отличается. В мелководном озере Виктория среднее содержание растворенных солей менее ОД г/л, Танганьика — 0,4, Эдуард — 0,8, Киву—1 г/л. Содержание свободного кислорода в водах рек и озер изменяется от 0,3 мг/л у дна до 7,8 мг/л у поверхности.
Имеют место случаи накопления в глубинных водах озер метана (Киву) и сероводорода (Эдуард), газы удерживаются высоким давлением воды (Д. Е. Хатчинсон, 1969).
Растительность лесных ландшафтов тропиков в геохимическом плане изучена слабо. Видовой состав леса разнообразен, в результате возникает сложность в выявлении закономерностей биологического круговорота. С переходом от экваториального к тропическому лесу в связи с сезонным изменением внешних факторов миграции картина биологического круговорота постепенно изменяется.
Рост и развитие растений в тропиках определяются многими условиями. Гиганты экваториального леса обязаны поступлению достаточного количества света, тепла, влаги, элементов питания для построения органических соединений. Карликовые деревья, как форма приспособления растительных организмов в процессе межвидовой борьбы и за источники питания, довольствуются меньшим количеством прямой солнечной радиации, потребляют меньше питания. В лесах, формирующихся на бедных питательными элементами почвах, достаточное количество тепла, влаги и света в течение года не способствует росту пышной растительности.
Образование органического вещества идет активно в течение года, и биомасса тропического леса зависит от особенностей сочетания внешних условий произрастания, наличия элементов питания, возраста пород.
Формы и состав растительного мира тропиков в некоторой степени отличаются от однотипных соединений в растениях умеренного пояса. Например, у злаков группы Graminees sacchariferes (тропики) запасные углеводы накапливаются в форме крахмала, а у злаков группы Graminees levilferes (умеренный пояс) — в форме фрук-тозана. Растворимость углеводов у растений тропиков ниже (3,5—4,5%), в растениях умеренного пояса выше (8,5—9,5%) (Е. С. Белюченко, 1978). Тропические растения содержат меньше протеина в гумидном климате, но больше клетчатки. У них накапливается больше насыщенных (твердых) жирных кислот типа стеариновой, а растения умеренного пояса содержат больше ненасыщенных (жидких) кислот. В растениях лесных ландшафтов тропиков накапливается больше углеводов и мало белков. С избытком углеводов в пище местного населения связывают болезнь спру—угнетение функций пищеварительных желез. Дефицит белков животного происхождения приводит к заболеванию квашиоркор. Эта болезнь, которая встречается преимущественно у детей, выражается в истощении организма, отставании в росте, общем понижении физического развития. В растениях лесных ландшафтов тропиков образуются органические соединения, не свойственные растениям умеренного пояса, например углевод маннан в плодах пальм, каучук в гевее.
Биомасса во влажных тропических, субэкваториальных и экваториальных лесах колеблется от 1300 (мантры) до 17 000 ц/га (гилей). В структуре биомассы на зеленую часть приходится 9%, многолетнюю надземную — 72, на корни — 19%. Прирост изменяется от 65 до 1000 ц/га, т. е. от среднего до высокого; зеленая и многолетняя надземная часть близки и соответственно составляют 47 и 42%. Химические элементы в структуре биомассы распределены неравномерно; около 48% приходится на многолетнюю надземную часть и 26%—на зеленую часть и корни. Высокая концентрация элементов в многолетней надземной части говорит о наличии ярко выраженного биологического барьера (Л. Е. Родии, Н. И. Базилевич, 1965).
Содержание зольных элементов в биомассе незначительно и составляет 81,41 ц/га, или 1,5%. Ежегодный прирост аккумулирует в среднем до 16 ц/га зольных элементов, или 4,7%. Содержание азота в приросте меньше суммы зольных элементов в четыре раза. Таким образом, в молодых органах растений на единицу веса аккумулируется больше химических элементов, чем в многолетних. Зольные элементы в биомассе распределяются по базипетальному типу: на долю листьев в среднем приходится до 3%, корней — 2,23, древесины и веток— 1%. Зольные элементы и азот образуют следующий ряд потребления химических элементов: N>Si> >Ca>K>Mg>S>Al>P>Fe>Mn>Na>Cl. Высокое содержание кремния коррелирует с повышенным содержанием его в речных и почвеино-грунтовых водах. По А. И. Перельману, растения усваивают за год в 4— 4,5.раза больше элементов, чем их выносится с ионным стоком. Это обстоятельство способствует удержанию подвижных мигрантов и предохраняет ландшафт от регрессивного развития,
Опад зеленой части протекает постепенно в течение всего года; он активно минерализуется микроорганизмами, поэтому подстилка не накапливается в большом количестве (20—100 ц/га), гумуса образуется мало. Интенсивность биологического круговорота высокая и составляет около 0,1. Ежегодно опадает 275 ц/га отмирающих растительных остатков, что составляет 5% от биомассы. С опадом возвращается на 5 ц/га меньше зольных элементов, чем потребляется приростом, азота в 2 раза меньше; зольные элементы и азот образуют следующий ряд: Si>N>Ca>Mg>Fe>Al>K>S>Mn> >P>Na>Cl. В золе онада на долю кремния приходится 50—60%, азота—17—25, щелочные и щелочноземельные основания в сумме составляют всего лишь 20—25%. Такое же примерно соотношение зольных элементов в биомассе древесной растительности, поэтому тип химизма растений лесных ландшафтов жаркого пояса азотно-кремниевый. Освобождающиеся при минерализации опада щелочные и щелочноземельные элементы активно выносятся из почвы, поэтому образующиеся в избытке органические и минеральные кислоты не могут быть полностью нейтрализованы, что приводит к формированию в ландшафте кислой среды.
Химические элементы поступают в почву также в результате смыва с поверхности листьев атмосферными осадками К, Na, CI, S (до 80%) в большем количестве, сульфаты (до 11%), Са, Mg (около 3—5%), фосфор (меньше 1%) —в меньшем.
Прикладные аспекты геохимии ландшафта
Миграция элементов в сельскохозяйственных ландшафтах. Растения лесных ландшафтов тропиков приспособлены к росту и развитию в условиях 12-часового светового дня в течение всего года. В спектре световых лучей в связи с повышенным содержанием озона преобладают тепловые инфракрасные лучи и мало ультрафиолетовых. Растения делятся на тене- и светолюбивые, поскольку поток прямой солнечной радиации в тропиках весьма интенсивен. Чтобы вызвать постоянное затенение почвы, в агротехнике используется подсев однолетних культур, проводится выращивание покровных растений в плантациях многолетних культур, а также быстро растущих деревьев; мульчирование, которое препятствует испарению влаги и защищает почву от прямого теплового облучения.
При выращивании сельскохозяйственных культур предусматриваются особенности миграции химических элементов в агроценозах как и в любом типе ландшафта. Растения тропиков усваивают неодинаковое количество химических элементов. Арахису нужно достаточное количество кальция; рис, просо, сахарный тростник потребляют много азота; банан, табак, кофейное дерево, сахарный тростник отзывчивы на внесение калийных удобрений. Существуют различия в потреблении элементов питания растениями и по фазам развития. Сахарный тростник усваивает в начале развития много азота и калия, к периоду созревания — фосфора. Масличные культуры используют фосфор равномерно в течение всеТо вегетационного периода.
На обрабатываемой почве хорошо развиваются нитрифицирующие бактерии, они переводят соединения азота в усвояемую нитратную форму. Нитраты перехватываются корневой системой, поэтому в круговороте почва — растение — опад — почва потери азота незначительны. Однако условия для развития азотфиксирую-hi их бактерий неблагоприятны из-за недостатка в почве кальция. В то же время при сжигании органических остатков азот из органических соединений переходит в золу, частично в газообразное состояние. Это приводит к потере азота из почвы в результате высокой миграционной способности соединений в обоих случаях. Запасы азота пополняются за счет сидеральных культур. На один гектар пашни мышей дает 831 кг азота, 861 кг калия, слоновая трава — 775 кг/га азота, 970 кг калия, 186 кг кальция, 108 кг магния. Отсюда следует, что зеленое удобрение дает меньше азота, чем калия. Запасы азота в почве рекомендуется восстанавливать внесением мочевины, содержащей до 46% азота. При повышенной влажности воздуха она мало гигроскопична. Минеральные азотные удобрения вносят в дозах 1—2 ц/га, что обусловлено высокой растворимостью азотных удобрений и потерей их из почвы. Такая доза обеспечивает потребность растений только на один сезон.
Органические и минеральные соединения фосфора в почвах тропиков трудно усваиваются растениями, так как слабо растворяются в воде. Наиболее распространены соединения ортофосфорной кислоты с алюминием, железом, кальцием. Кислая реакция ускоряет растворение фосфатов. Фосфат-ионы, сорбированные коллоидными частицами/становятся доступными для растений. Исключение составляют коллоидные частицы оксидов железа, которые прочно удерживают фосфор при нейтральной или щелочной реакции. Слоновая трава как зеленое удобрение дает до 22 кг/га фосфора, т. е. в 35 раз меньше, чем азота. В компосте из торфа содержится от 0 до 3% фосфора. Как удобрение хорошо зарекомендовал себя томас-шлак, поскольку фосфор освобождается из него постепенно. Фосфоритов вносят в почву 10—30 ц/га, суперфосфата — 5—8 ц/га.
В почвах тропиков калия мало, но его содержание возрастает с увеличением засушливости и с повышением почвенной кислотности, а также в почвах с преобладанием гидрослюды. В связи с активным гидролизом первичных минералов калий освобождается быстрее, чем в умеренном поясе. Много калия поступает в почву с зелеными сидератами, однако его мало в компосте из торфа (0—0,2%). Эффективно применение негигроскопического сульфата калия в дозе 1,5—3 ц/га; сильвинита и каинита вносят в почву в дозе до 4—6 ц/га, калийной соли около 1—3 ц/га.
В настоящее время научно обоснованная система внесения основных минеральных удобрений (NPK) находится в стадии разработки. Рекомендации по внесению NPK по разным странам с одинаковыми условиями отличаются в силу традиций. Например, в Сенегале под арахис рекомендуется вносить азотно-фосфорно-калий-ные удобрения в соотношении 14:7:7, в Мали — 6 : 20 : 10, в Западной Африке — 6 : 20 : 0.
Из других химических элементов, необходимых для роста и развития растений, в почвах тропиков с гумид-ным климатом недостает кальция, магния, из микроэлементов— бора и серы. Сидеральные культуры дают до 200 кг/га кальция и до 100 кг/га магния. Кальций используется как удобрение для арахиса и земляного ореха, а также как средство понижения кислотности почвы. Культурные растения хорошо реагируют на внесение серы в почву в дозе 25 кг/га. Общее содержание железа в почвах высокое, однако окислительная обстановка препятствует его переходу в подвижную форму. Недостаток элементов В, Mn, Си, Zn, I, Br, Mo, Со ощущают растения на торфяных, легких минеральных, а также на древних почвах. Учитывая интенсивный промывной режим, удобрения в почву следует вносить часто и небольшими дозами.
В связи с тем, что вегетационный период длится в течение года и за это время снимается несколько урожаев сельскохозяйственных культур, отчуждение с урожаем химических элементов достигает значительной величины. Вынос питательных элементов с урожаем дает примерное представление о том, какую дозу минеральных удобрений необходимо вносить ежегодно.
Геохимия ландшафта и здравоохранение. Биогеохимические эндемии в лесных ландшафтах тропиков изучены недостаточно. Имеются сведения о низком содержании многих элементов питания растений, что проявляется в изменении цвета листьев, сосудов в стволе и ветвях, анормальном развитии или замедлении роста. Особенно часто страдают от недостатка элементов питания цитрусовые и масличная пальма. Дефицит элементов питания растений отражается на росте и продуктивности животных.
Дата добавления: 2015-06-27; просмотров: 1970;