Регуляция дыхания.
Как и все системы в организме, дыхание регулируется двумя основными механизмами – нервным и гуморальным.
Основой нервной регуляции является реализация рефлекса Геринга-Бреера, который по сути, состоит из серии последовательно в процессе дыхания сменяющихся друг друга рефлексов, подобно описанных различных учебниках по физиологии. Здесь отметим, что все рефлексы могут быть объединены как один, суть которого заключается в следующем: вдох, выдох стимулирует вдох.
Смене дыхательных фаз способствуют сигналы, поступающие от механорецепторов легких по афферентным волокнам блуждающих нервов. Импульсы, поступающие от рецепторов легких, обеспечивают смену вдоха на выдох и смену выдоха вдохом (рис.7).
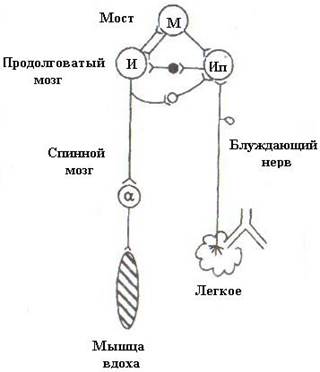
Рис.7. Схема, отражающая основные процессы саморегуляции вдоха и выдоха.
И – совокупность инспираторных нейронов, обеспечивающих вдох; Ип – инспираторные поздние нейроны, прерывающие вдох: светлые – возбуждающие, темные – тормозящие.
В эпителиальном и субэпителиальном слоях всех воздухоносных путей, а также в области корней легких расположены так называемые ирритантные рецепторы, которые обладают одновременно свойствами механо- и хеморецепторов. Они раздражаются при сильных изменениях объема легких. Ирритантные рецепторы возбуждаются также под действием пылевых частиц, паров едких веществ и некоторых биологически активных веществ, например, гистамина. Однако для регуляции смены вдоха и выдоха большее значение имеют рецепторы чувствительные к растяжению легких (механическое раздражение).
В продолговатом мозге нейроны, ответственные за ритмическую смену актов вдоха и выдоха, формируют несколько ядер дорсальной и вентральной групп, среди которых последняя имеет большее значение в реализации рефлекса Геринга-Бреера. Условно все ядра вентральной и дорсальной групп можно объединить общим названием дыхательного центра (ДЦ). Нейроны дыхательного центра продолговатого мозга как бы разделены на две группы. Одна группа нейронов дает волокна к мышцам, которые обеспечивают вдох, эта группа нейронов получила название инспираторных нейронов (инспираторный центр, ИЦ), т.е. центр вдоха. Другая же группа нейронов, отдающих волокна к внутренним межреберным и межхрящевым мышцам, получила название экспираторных нейронов (экспираторный центр, ЭЦ), т.е. центр выдоха. Нейроны экспираторного и инспираторного отделов дыхательного центра продолговатого мозга обладают различной возбудимостью и лабильностью. Возбудимость инспираторного отдела выше. Кроме того ИЦ обладает выраженной автоматией.
Вдох начинается с возбуждения ИЦ, которое во многом обеспечивается автоматическими процессами в нем. Импульсация по нисходящей поступает по мотонейронам спинного мозга, аксоны, которых составляют диафрагмальный, наружные межреберные и межхрящевые нервы, иннервирующие основные мышцы вдоха. Сокращение этих мышц увеличивает размер грудной клетки, воздух поступает в альвеолы, растягивая их. Чем глубже происходит вдох, тем в большей степени активируются рецепторы легкого. Частота афферентации от них нарастает, направляясь в ЭЦ, который возбуждается. Возбуждение ЭЦ наводит торможение на ИЦ, моторная импульсация от него к мышцам вдоха прекращается, которые расслабляются. Происходит пассивный, под действием силы тяжести, выдох. Т.о. вдох стимулирует выдох.
При выдохе паренхима легкого спадается, прекращается активация ее механорецепторов, значит, исчезает афферентация к ЭЦ. Возбуждение ЭЦ прекращается и он перестает наводить торможение на ИЦ. В последнем нарастают автоматические процессы, и он возбуждается. Начинается новый акт вдоха, т.е. выдох стимулирует вдох.
Конечно, формирования стволовыми структурами паттерна дыхания, здесь описанного Люмсденом (1920), приводится здесь в упрощенном виде. В действительности дыхательные нейроны продолговатого мозга формируют несколько вентральных и дорсальных групп, ответственных за генерацию различного типа моторной импульсации в разные моменты (начало – конец), как вдоха, так и выдоха. Представляется нецелесообразным подробное изложение в настоящем издании современных представлений о механизмах дыхательного ритмогенеза. Подчеркнем лишь, что два основных свойства дыхательного центра, обеспечивающих реализацию рефлекса Геринга-Бреера – это автоматия и реципрокность. Способность самовозбуждаться присутствует не только в инспираторных нейронах, как это описано выше, но и в ЭЦ. Кроме того, как ЭЦ, способен наводить торможение на ИЦ, так и наоборот. Между этими двумя группами дыхательных нейронов существуют антогонистические (реципрокные) взаимоотношения.
Кроме того, обратим внимание, что дыхательный центр, расположенный в продолговатом мозге, способен формировать ритм внешнего дыхания посредством нервного (рефлекторного) механизма. Однако, известно, что интенсивность дыхания в большей степени зависит и от гуморальных факторов, например, кислотности крови, а также может быть изменена произвольно.
Значительный вклад в изучение данных механизмов был сделан отечественным физиологом Н.А. Миславским, на основе работ которого можно ввести понятие центрального механизма регуляции дыхания (рис.8).
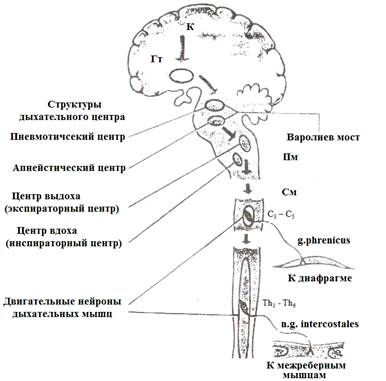
Рис.8. Дыхательный центр (его компоненты) и эфферентные нервы.
К – кора; Гт – гипоталамус; Пм – продолговатый мозг; См – спинной мозг; Тh1-Тh6 – грудной отдел спинного мозга; С3-С5 – шейный отдел спинного мозга.
Центральный механизм регуляции дыхания (ЦМРД) – это вся совокупность ядер головного мозга, участвующих в формирования ритма и глубины дыхательных движений. Основными элементами ЦМРД являются ДЦ продолговатого мозга, пневмотоксический центр (ПТЦ) среднего мозга, кора больших полушарий (КГМ).
Дыхательный центр продолговатого мозга испытывает влияния со стороны вышележащих отделов ЦНС. Так, например, в передней части варолиева моста расположен ПТЦ, который способствует периодической деятельности дыхательного центра, он увеличивает скорость развития инспираторной активности, повышает возбудимость механизмов выключения вдоха, ускоряет наступление следующей инспирации. Другими словами, ПТЦ интенсивно обменивается возбуждающей и тормозной импульсацией с инспираторными и экспираторными нейронами продолговатого мозга. ПТЦ повышает или понижает возбудимость ДЦ, изменяя, тем самым, внешнее дыхание.
По современным представлениям возбуждение клеток инспираторного отдела продолговатого мозга активирует деятельность апноэстического и пневмотаксического центров. Апноэстический центр тормозит активность экспираторных нейронов, пневмотаксический – возбуждает. По мере усиления возбуждения инспираторных нейронов под влиянием импульсации от механо- и хеморецепторов усиливается активность пневмотаксического центра. Возбуждающие влияния на экспираторные нейроны со стороны этого центра к концу фазы вдоха становятся преобладающими над тормозными, приходящими со стороны апноэстического центра. Это приводит к возбуждению экспираторных нейронов, оказывающих тормозящие влияния на инспираторные клетки. Вдох тормозится, начинается выдох.
Существует самостоятельный механизм торможения вдоха и на уровне продолговатого мозга. К этому механизму относят специальные нейроны (I бета), возбуждаемые импульсами от механорецепторов растяжения легких и инспираторно-тормозные нейроны, возбуждаемые активностью нейронов I бета. Таким образом, при увеличении импульсации от механорецепторов легких увеличивается активность I бета нейронов, что в определенный момент времени (к концу фазы вдоха) вызывает возбуждение инспираторно-тормозных нейронов. Их активность тормозит работу инспираторных нейронов. Вдох сменяется выдохом.
Активность ПТЦ, зависит от многих факторов:
Во-первых, ПТЦ получает афферентацию от различны органов и систем организма: рецепторов паренхимы легкого, сосудистых рефлексогенных зон, других рецептивных полей. Во-вторых, ПТЦ имеет собственные центральные хеморецепторы, чувствительные к изменению кислотности и газового состава ликвора. Таким образом, гуморальная регуляция внешнего дыхания осуществляется во многом благодаря ПТЦ. В-третьих, ПТЦ, находится в тесном взаимодействии с КГМ и под ее контролем, чем обеспечивается произвольная регуляция дыхания.
Если произвести перерезку путей, соединяющих ПТЦ, с КГМ, то внешнее дыхание практически не изменится. У животного в полном объеме сохраниться возможность адаптации интенсивности дыхания к меняющимся условиям существования, которая будет осуществляться по безусловно рефлекторному типу с участием ПТЦ и ДЦ. Однако, произвольная регуляция окажется невозможной, например, будет происходить задержки дыхания при погружении головы в воду.
Если затем произвести перерезку ствола мозга ниже мезенцефального отдела (средний мозг), выключив, тем самым, ПТЦ, то внешнее дыхание сохранится, но изменится значительно (рис.9).
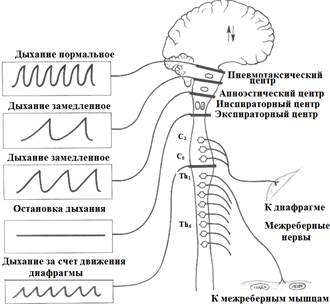
А В
Рис.9. Влияние на дыхание перерезок на разных уровнях головного и спинного мозга. А – характер дыхательных движений, В – уровни перерезок.
Оно будет состоять из чередующихся фаз глубоких вдоха и выдоха, т.е. будет реализовываться только в соответствии с рефлексом Геринга-Бреера. При этом гуморальная регуляция окажется практически невозможной, например, закисление крови не будет приводить к увеличению глубины дыхания. Наконец, полное отсечение головного мозга от спинного приводит к остановке дыхания.
В регуляции дыхания большое значение имеют центры гипоталамуса. Под влиянием центров гипоталамуса происходит усиление дыхания, например, при болевых раздражениях, при эмоциональном возбуждении, при физической нагрузке.
Говоря о гуморальной регуляции внешнего дыхания, следует отметить, что деятельность дыхательного центра в значительной степени зависит от напряжения газов в крови и концентрации в ней водородных ионов. Ведущее значение в определении величины легочной вентиляции имеет напряжение углекислого газа в артериальной крови, оно как бы создает запрос на нужную величину вентиляции альвеол.
Содержание кислорода и особенно углекислого газа поддерживается на относительно постоянном уровне. Нормальное содержание кислорода в организме называется нормоксия, недостаток кислорода в организме и тканях – гипоксия, а недостаток кислорода в крови – гипоксиемия. Увеличение напряжения кислорода в крови называется гипероксия. Нормальное содержание углекислого газа в крови называется нормокапния, повышение содержания углекислого газа – гиперкапния, а снижение его содержания – гипокапния.
Нормальное дыхание в состоянии покоя называется эйпноэ. Гиперкапния, а также снижение величины рН крови (ацидоз) сопровождаются увеличением вентиляции легких - гиперпноэ, что приводит к выделению из организма избытка углекислого газа, увеличение вентиляции легких происходит за счет увеличения глубины и частоты дыхания.
Гипероксия, гипокапния и повышение уровня рН крови приводит к уменьшению вентиляции легких, а затем и к остановке дыхания - апноэ.
Деятельность ДЦ зависит от состава крови, поступающей в мозг по общим сонным артериям. В 1901 г. это было показано Л. Фредериком опытах с перекрестным кровообращения. У двух собак, находившихся под наркозом, перерезали и соединяли перекрестно сонные артерии и яремные вены. При этом голова первой собаки снабжалась кровью второй собаки, и наоборот. Если у одной из собак, например, первой, перекрывали трахею и таким путем вызывали асфиксию, то гиперпноэ развивалось у второй собаки. У первой же собаки, несмотря на увеличение в артериальной крови напряжение СО2 и снижение О2, развивалось апноэ, так как в ее сонную артерию поступала кровь второй собаки, у которой в результате гипервентиляции снижалось напряжение СО2 в артериальной крови (рис.10).
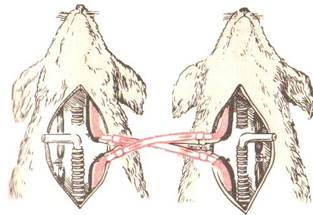
Рис.10. Опыт с перекрестным кровообращением (по Л. Фредерику).
Двуокись углерода, водородные ионы и умеренная гипоксия вызывают усиление дыхания. Эти факторы усиливают деятельность дыхательного центра, оказывая влияние на периферические и центральные хеморецепторы, регулирующие дыхание.
Роль рефлексогенных зон в регуляции дыхания.
Хеморецепторы, чувствительные к увеличению напряжения углекислого газа и к снижению напряжения кислорода находятся в каротидных синусах и дуге аорты. Большее значение для регуляции дыхания имеют каротидные хеморецепторы. При нормальном содержании кислорода в артериальной крови в афферентных нервных волокнах, отходящих от каротидных телец, регистрируются импульсы. При снижении напряжения кислорода частота импульсов возрастает особенно значительно, т.к. гипоксия оказывает на артериальные хеморецепторы стимулирующее влияние. Кроме того, афферентные влияния с каротидных телец усиливаются при повышении в артериальной крови напряжения углекислого газа и концентрации водородных ионов. Хеморецепторы каротидных телец, информируют дыхательный центр о напряжении О2 и СО2 в крови, которая направляется к мозгу.
Дыхание зависит от рефлекторных влияний с сосудистых рефлексогенных зоне, и в частности, с барорецепторов зоны позвоночных артерий (ЗПА). В частности, ЗПА вызывает сочетанные изменения дыхания и системного артериального давления.
Центральные хеморецепторы расположены в продолговатом мозге, они постоянно стимулируются водородными ионами, находящимися в спинномозговой жидкости. Перфузия этой области мозга раствором со сниженным рН резко усиливает дыхание, а при высоком рН дыхание ослабевает, вплоть до апноэ. То же происходит при охлаждении или обработке этой поверхности продолговатого мозга анестетиками. Центральные хеморецепторы, оказывая сильное влияние на деятельность дыхательного центра, существенно изменяют вентиляцию легких.
Центральные хеморецепторы реагируют на изменение напряжения СО2 в артериальной крови позже, чем периферические хеморецепторы, так как для диффузии СО2 из крови в спинномозговую жидкость и далее в ткань мозга необходимо больше времени. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз – тормозят центральные хеморецепторы.
Импульсы, поступающие от центральных и периферических хеморецепторов, являются необходимым условием периодической активности нейронов дыхательного центра и соответствия вентиляции легких газовому составу крови.
Своеобразие функции внешнего дыхания состоит в том, что она одновременно и автоматическая, и произвольно управляемая.
Дата добавления: 2015-06-22; просмотров: 3362;