Структура нуклеиновых кислот
При гидролизе нуклеиновых кислот образуются пуриновые (аденин, гуанин) и пиримидиновые (тимин, цитозин, урацил) основания, углевод (рибоза или дезоксирибоза) и фосфорная кислота. Отличиями в строении ДНК и РНК являются: в ДНК углевод дезоксирибоза, в РНК – рибоза; в ДНК содержится тимин, в РНК – урацил. В составе ДНК и РНК открыты так называемые минорные азотистые основания, на долю которых приходится до 10% всех нуклеотидов.
Азотистое основание соединено с углеводным компонентом гликозидной связью. В образовании гликозидной связи в пуриновых нуклеотидах принимает участие N-9 пурина и С-1' пентозы, а в пиримидиновых нуклеотидах – N-1 пиримидина и С-1' пентозы.
Для того, чтобы отличить углеродные атомы рибозы или дезоксирибозы от углеродных атомов, входящих в состав пуриновых или пиримидиновых оснований, первые принято обозначать символом «штрих».
К 5’-углеродному атому углевода присоединяется остаток фосфорной кислоты (от 1 до 3) и образуются нуклеотиды.
Нуклеозид состоит из азотистого основания и углеводного компонента. Нуклеотид – нуклеозид + фосфорная кислота.
Основные функции ДНК по А. Ленинджеpу: 1) хpанение запаса генетической инфоpмации, необходимой для кодиpования стpуктуpы всех белков и всех РНК каждого вида оpганизма; 2) pегуляция во вpемени и пpостpанстве биосинтеза компонентов клеток и тканей; 3) опpеделение деятельности оpганизма в течение его жизненного цикла; 4) обеспечение индивидуальности данного оpганизма.
Различают следующие основные виды ДНК: 1) ядерные (хромосомные) ДНК; 2) ДНК плазмид; 3) ДНК хлоропластов; 4) ДНК митохондрий; 5) ДНК вирусов.
Ядерная ДНКлокализована в ядре эукариотической клетки. Аналогом ядерной ДНК у бактерий служит генофор, или нуклеоид, который представляет собой кольцевидно замкнутую ДНК, не отделенную от цитоплазмы мембраной. Нередко генофор называют бактериальной хромосомой.
Молекулы ядерных ДНК содержат основной объем информации обо всех наследственных признаках организма. Их функция – хранение этой информации, обеспечение ее экспрессии и рекомбинации, а также воспроизводство при делении клетки и передача последующим поколениям организма.
ДНК плазмид – это обычно кольцевидно замкнутые молекулы ДНК небольшого размера, находящиеся в цитоплазме. Несмотря на небольшой объем содержащейся в них наследственной информации, ДНК плазмид играют важную роль в явлениях наследственности и изменчивости организмов, прежде всего бактерий и грибов. Плазмидные ДНК – это относительно автономные структуры: они способны реплицироваться независимо от ядерной ДНК, а также переходить из одной клетки в другую в процессе их контактов. Эту способность плазмидных ДНК широко используют инженеры-генетики для переноса наследственной информации из одних организмов в другие.
ДНК хлоропластов находятся в пластидах растений. Они несут информацию о белках и РНК, необходимых для нормальной работы фотосинтетического аппарата растительной клетки. Поскольку хлоропласты растений обладают автономным аппаратом для синтеза белка, то роль этих ДНК отчасти аналогична функциям ядерной ДНК.
ДНК митохондрий, подобно ДНК хлоропластов, обеспечивают автономный синтез белка, который протекает в митохондриях. Несмотря на известную автономность, функционирование ДНК хлоропластов и ДНК митохондрий четко координировано с деятельностью ядерной ДНК. Эти виды ДНК в совокупности образуют единый наследственный аппарат эукариотической клетки
ДНК вирусов. Значительная часть вирусов бактерий (бактериофагов), животных и человека принадлежит к ДНК-содержащим вирусам. Например, возбудители оспы человека, герпеса, гепатита В – ДНК-содержащие вирусы. Гораздо меньше их (не более 3%) среди вирусов растений и грибов.
Вирусные ДНК несут относительно небольшой объем наследственной информации (от 10 до 150 генов), обеспечивающей циркуляцию этих паразитов в природе, в частности инфицирование клетки-хозяина.
Подобно плазмидным ДНК, вирусные ДНК обладают способностью «путешествовать» между клетками и встраиваться (интегрироваться) в их ДНК. Поэтому ДНК-содержащие вирусы находят применение в качестве носителей наследственной информации в генно-инженерных разработках.
Нуклеотиды в нуклеиновых кислотах связаны 3’,5’-фосфодиэфирными связями, которая возникает между 3’-ОН группой углевода одного нуклеотида и 5’-ОН группой углевода другого нуклеотида.
РНК – одноцепочечная молекула. Однако при наличии в цепи РНК участков с комплементарной последовательностью единичная цепь РНК способна сворачиваться с образованием так называемых «шпилек», структур, имеющих двуспиральные характеристики.
Для понимания ряда особенностей первичной структуры ДНК важное значение имеют закономерности количественного содержания азотистых оснований, установленные впервые Э.Чаргаффом в 1949 году и названные правилами Чаргаффа:
1. Количество пуриновых оснований равно количеству пиримидиновых оснований.
2. Количество аденина и цитозина равно количеству гуанина и Тимина.
3. Количество аденина равно количеству тимина, а количество гуанина равно количеству цитозина.
4. Коэффициент специфичности, который отражает 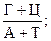 для эукариот этот коэффициент ниже единицы (0,54 – 0,94), для прокариот – выше единицы.
для эукариот этот коэффициент ниже единицы (0,54 – 0,94), для прокариот – выше единицы.
В соответствии с моделью Дж. Вторичная структура ДНК
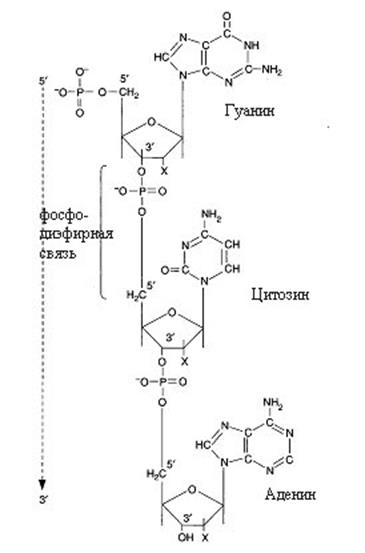
Уотсона и Ф.Крика, предложенной в 1953 г. на основании ряда аналитических данных, а также рентгеноструктурного анализа, молекула ДНК состоит из 2-х цепей, образуя правовращающую спираль, в которой обе полинуклеотидные цепи закручены вокруг одной и той же оси. Удерживаются полинуклеотидные цепи водородными связями, образующимися между комплементарными азотистыми основаниями: между А и Т – две водородныесвязи, Ц и Г – три водородные связи. Азотистые основания расположены внутри спирали, а фосфорные остатки и углеводные компоненты – снаружи. Кроме водородных связей в стабилизации молекулы ДНК принимают участие силы гидрофобного («стэкинг») взаимодействия, образующегося между плоскостями оснований внутри данной цепи (рис. 12.1).
Обе цепи в молекуле ДНК имеют противоположную полярность (антипараллельны). Это означает, что одна цепь имеет направление 5'→ 3', а другая 3'→5'. Подобная направленность цепей имеет важное биологическое значение при репликации и транскрипции молекулы ДНК.
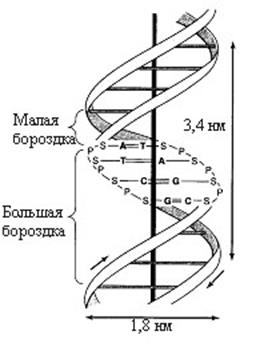
Рис. 12.1. Двойная спираль ДНК
Конфигурация двойной спирали ДНК меняется в зависимости от количественного содержания в ней воды и ионной силы окружающей среды. В настоящее время известно шесть форм ДНК (от А до Е и Z-форма). При физиологических условиях доминирующим структурным типом ДНК является В-форма. Расстояние между витками спирали (или шаг спирали) равно 3,4 нм. На этом участке укладывается 10 нуклеотидных остатков, размер одного нуклеотида составляет 0,34 нм, диаметр биспиральной молекулы 1,8 нм. В структуре ДНК различают большую и малую бороздки, закрученные вокруг оси молекулы параллельно фосфодиэфирному остову. В этих бороздках белки могут специфически взаимодействовать с определенными атомами нуклеиновых оснований.
Среди разнообразных конформаций ДНК различают линейную ДНК и кольцевидно замкнутую ДНК. Кольцевидная структура ДНК характерна для бактерий и некоторых вирусов.
ДНК в клетке имеет длину 1,74 м, поэтому понятно, что в ядре происходит упаковка ДНК. Третичная структура ДНК прокариот может образовываться в результате дополнительного скручивания в пространстве двуспиральной молекулы с образованием суперспирали или суперкольца. У высших организмов ДНК находится в хромосомах, которые при митозе видны в световой микроскоп. В каждой хромосоме находится молекула ДНК, которая составляет основу хроматина. Хроматин – комплекс ДНК с РНК и белками (ДНК 30-45%, гистоны 30-50%, негистоновые белки 4-30%, РНК до 10%). Структурная организация хроматина такова, что позволяет использовать одну и ту же генетическую информацию ДНК по-разному в специализированных клетках. При этом основная часть хроматина не активна. Активный хроматин составляет в разных клетках от 2 до 11%. Различают несколько уровней укладки (компактизации) ДНК.
1) Нуклеосомы. В электронном микроскопе изображение хроматина напоминает бусы: шаровидные утолщения размером около 10 нм, разделенные перемычками. Каждая нуклеосома содержит отрезок двуспиральной ДНК, равный по протяженности примерно 145-150 парам оснований, обернутый в 1,5 оборота вокруг ядра, состоящего из гистонов (2Н1, 2Н2, 2Н3 и 2Н4). Свободные от контакта с белками участки ДНК называют линкерными (или связующими). Их длина варьирует в зависимости от типа клеток (от 15 дог 100 нм). Линкерные участки ДНК либо свободны, либо контактируют с гистонами Н1. Степень компактизации в 5 раз. Примерно 90% ДНК входит в состав нуклеосом, 10% содержится в перемычках между нуклеосомами. Считают, что нуклеосомы содержат фрагменты «молчащего» хроматина, а перемычки – активного. При развертывании весь хроматин активный. Межнуклеосомный разрыв ДНК - способ запрограммированной гибели клеток – апоптоз. Из нуклеосом образуются фибриллы (рис. 12.2).
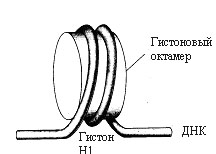
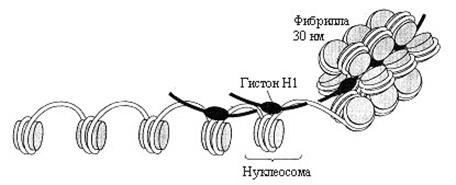
Рис. 12.2. Нуклеосомы
2) Фибриллы толщиной 10 нм состоят из ряда нуклеосом, касающихся друг друга своими краями и ориентированных плоскими поверхностями вдоль оси фибриллы. Эта структура называется соленоид. Компактизация в 40 раз.
3) Фибриллы скручиваются в спираль, на виток которых приходится 6-7 нуклеосом. В результате образуется хроматиновое волокно диаметром 30 нм.
4) Для того, чтобы образовалась митотическая хромосома нормального размера, волокно диаметром 30 нм должно подвергнуться дополнительной компактизации с результирующим уменьшением длины еще в 100 раз.
Транскрипционно-неактивный хроматин (гетерохроматин) плотно упакован и поэтому соответствующие области интенсивно окрашиваются. Участки транскрипционно-активного хроматина (эухроматина) имеют более слабую окраску.
Двухцепочечные стpуктуpы ДНК пpи нагpевании, пpи экстpемальных значениях pН, обpаботке мочевиной и дp. могут пеpеходить в фоpму неупоpядоченных клубков - денатуpация ДНК. Молекулы нуклеиновых кислот максимально поглощают ультpафиолет пpи 260 нм за счет поглощения азотистых оснований. Раствоp нативной ДНК имеет пpи 260 нм оптическую плотность на 40% ниже оптической плотности смеси нуклеотидов. Это явление называют гипеpхpомным эффектом. О денатуpации ДНК судят по увеличению поглощения ультрафиолета при 260 нм. Пpи нагpевании поглощение пpи 260 нм возpастает в узком диапазоне темпеpатуp - точка плавления (80-85 ºС): Е260 ДНК < Е260 смеси нуклеотидов на 40%. Денатуpация обpатима, если остались спиpализованные участки ДНК. Восстановление стpуктуpы ДНК после удаления денатуpизующего фактоpа (за счет комплементаpного спаpивания оснований нуклеотидов) называется pенативацией ДНК (отжиг ДНК). На явлении денатуpации-pенативации основан метод гибpидизации. Известны гибpидные двухцепочечные молекулы: - ДНК-РНК, обpазуются как пpомежуточные фоpмы пpи действии обpатной тpанскpиптазы; - ДНК-РНК на коpоткое вpемя в пpоцессе тpанскpипции ДНК. В проценссе трансляции и транскрипции происходит разделение цепей ДНК.
Дата добавления: 2015-06-12; просмотров: 2764;