Обмен нуклеотидов
Пуриновые азотистые основания, образующиеся в процессе переваривания нуклеиновых кислот в кишечнике, в дальнейшем практически не используются. Поэтому их синтез осуществляется de novo из низкомолекулярных предшественников, продуктов обмена углеводов и белков.
1. Для биосинтеза пуринового кольца необходимы следующие предшественники: N1 образуется из аспартата; С2 – из углерода N10-формил-ТГФК; N3 и N9 происходят из амидной группы глутамина; С4, С5 и N7 - из глицина; С6 – из СО2; С8 - N5,N10-метенил-ТГФК.
2. Основное место синтеза – печень. В эритроцитах, лейкоцитах и мозгу синтез не происходит.
3. Синтез пуриновых нуклеотидов начинается с промежуточного продукта пентозофосфатного пути – рибозо-5-фосфата, на который переносится пирофосфатная группа АТФ (схема). Фермент – фосфорибозилпирофосфатсинтетаза.
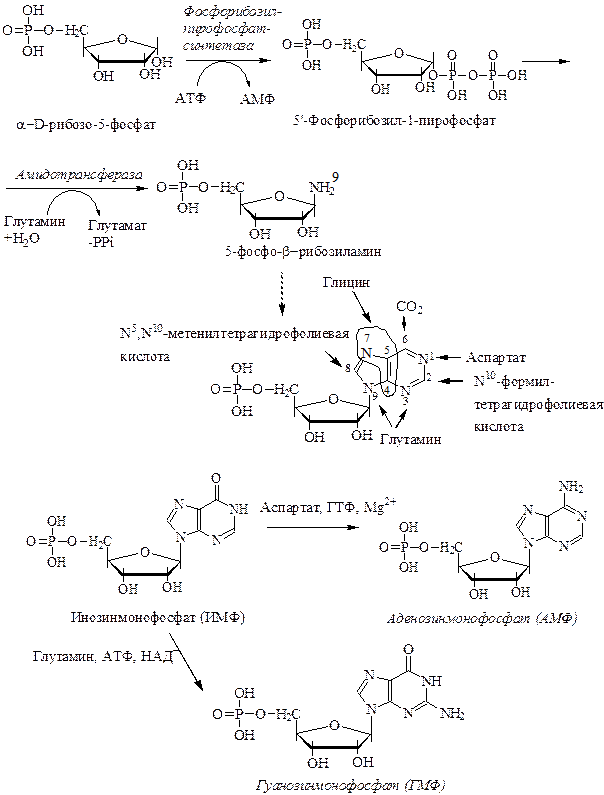
4. Образовавшийся 5-фосфорибозил-1-пирофосфат (ФРПФ) под действием амидотрансферазы взаимодействует с глутамином, который является донором NH2-группы и образуется β-5-фосфорибозиламин. При этой реакции происходит освобождение пирофосфата, глутаминовой кислоты и изменение конфигурации из α- в β-. Эта стадия является ключевой в синтезе пуринов.
5. Через ряд промежуточных соединений происходит синтез всего пуринового скелета и образуется первый нуклеотид – инозинмонофосфат (инозиновая кислота, ИМФ), которая является предшественником пуриновых нуклеотидов.
6. Синтез АМФ и ГМФ осуществляется из ИМФ. Для синтеза ГМФ необходимы НАД+, АТФ, глутамин и ферменты – ИМФ-дегидрогеназа и ГМФ-синтетаза. В ферментативном синтезе АМФ из ИМФ принимает участие аспарагиновая кислота, являющаяся донором NH2-группы, ГТФ в качестве источника энергии и ферменты аденилосукцинатсинтетаза и аденилосукцинатлиаза.
Превращение АМФ и ГМФ в соответствующие нуклеозидди- и трифосфаты протекает в две стадии при участии специфических нуклеозидмонофосфат – и нуклеозиддифосфаткиназ:
ГМФ + АТФ ↔ ГДФ + АФД
ГДФ + АТФ ↔ ГТФ + АДФ
Регуляция осуществляется по механизму обратной связи:
1. Синтез пуриновых нуклеотидов тормозится конечными продуктами по принципу обратной связи путем ингибирования амидотрансферазной реакции.
2. Избыток ГМФ в клетках аллостерически тормозит синтез ГМФ и не влияет на синтез АМФ и, наоборот, накопление АМФ ингибирует синтез АМФ и не ингибирует синтез ГМФ.
Ингибиторы синтеза пуриновых нуклеотидов: фолиевая кислота необходима дял синтеза пуриновых нуклеотидов. Сульфаниламидные препараты являются структурными аналогами парааминобензойной кислоты и ингибируют ее синтез у микроорганизмов. Сульфаниламиды не действуют на организм человека, поскольку фолиевая кислота не синтезируется у человека, а поступает с пищей.
Особенностью синтеза пиримидиновых нуклеотидов является то, что вначале происходит синтез пиримидинового основания, а затем остаток рибозы присоединяется к уже сформировавшемуся пиримидиновому кольцу.
1. Из глутамина, СО2 и АТФ под действием фермента карбамоилфосфатсинтетазыв цитозоле клеток синтезируется карбамоилфосфат (схема).
2. Карбамоилфосфат взаимодействует с аспартатом и образуется N-карбамоиласпарагиновая кислота. Фермент – аспартаттранскарбамоилаза.
3. N-карбамоиласпарагиновая кислота подвергается циклизации с отщеплением молекулы воды и дегидрированию. Синтезируется оротовая кислота.
4. Оротовая кислота реагирует с ФРПФ с образованием оротидин-5’-фосфата (ОМФ). Декарбоксилирование последнего приводит к образованию пиримидинового нуклеотида – УМФ.
5. Превращение УМФ в УДФ и УТФ осуществляется, как и в случае пуриновых нуклеотидов, путем фосфорилирования:
УМФ + АТФ ↔ УДФ + АФД
УДФ + АТФ ↔ УТФ + АДФ
6. Предшественником цитидиловых нуклеотидов является УТФ, который превращается в ЦТФ с участием фермента ЦТФ-синтетаза, АФТ, Mg2+ и глутамина.
7. Тимидиловые нуклеотиды входят в ДНК и содержат дезоксирибозу. Поэтому вначале происходит восстановление рибонуклеотидов у 2’-го атома углерода с использованием в качестве источника атомов водорода восстановленного термостабильного белка тиоредоксина, содержащего 2 свободные SH-группы.
8. Для синтеза тимидиловых нуклеотидов требуется также метилированное производное урацила – тимин. В клетках имеется фермент – тимидилатсинтаза, который катализирует метилирование дезоксиУМФ.
Донором метильной группы в тимидилатсинтазной реакции является N5,N10-метилен-ТГФК, которая превращается в дигидрофолиевую кислоту (ДГФК). Последняя восстанавливается до ТГФК под действием НАДФН2-зависимой дигидрофолатредуктазы. Ингибитор дигидрофолатредуктазы – метотрексат – используется как противоопухолевый препарат. Из образовавшегося ТМФ путем фосфотрансферазных реакций образуются dТДФ и dТТФ.
Дата добавления: 2015-06-12; просмотров: 3775;