рекет потенциалы.
 Жасушалы мембрананың қозу үрдісін Э. Бернштейн оқып-зерттеп, әртүрлі иондар үшін оның өтімділігін өлшеуге болатындығын айтқан. Бірақ ол осы мәселені оқып-зерттеуді аяқтауды үлгермеді. Шамамен 1939 ж. Коул мен Кертис (АҚШ) және Ходжкин мен Хаксли (Ұлыбритания) кальмардың үлкен аксонының қозбаған және қозған жасушаларының потенциалдарын өлшеді: тыныштық күйде потенциалдар айырмасы шамамен -80 мВ- қа (теріс) тең болатындығы, ал қозған кезде потенциалдардың максимал айырмасы +40мВ- қа (оң) шамамен тең болатындығы анықталды. Әрі қарай Ходжкин мен Хаксли мынадай гипотезаны тұжырымдады: қозу кезінде мембрананың әртүрлі иондар үшін өтімділігі өзгереді: оның негізінен калийді жақсы өткізгіштігі натрийдің өткізгіштігіне ауысады, демек натрий иондарының жасуша ішіне ағыны артады.
Жасушалы мембрананың қозу үрдісін Э. Бернштейн оқып-зерттеп, әртүрлі иондар үшін оның өтімділігін өлшеуге болатындығын айтқан. Бірақ ол осы мәселені оқып-зерттеуді аяқтауды үлгермеді. Шамамен 1939 ж. Коул мен Кертис (АҚШ) және Ходжкин мен Хаксли (Ұлыбритания) кальмардың үлкен аксонының қозбаған және қозған жасушаларының потенциалдарын өлшеді: тыныштық күйде потенциалдар айырмасы шамамен -80 мВ- қа (теріс) тең болатындығы, ал қозған кезде потенциалдардың максимал айырмасы +40мВ- қа (оң) шамамен тең болатындығы анықталды. Әрі қарай Ходжкин мен Хаксли мынадай гипотезаны тұжырымдады: қозу кезінде мембрананың әртүрлі иондар үшін өтімділігі өзгереді: оның негізінен калийді жақсы өткізгіштігі натрийдің өткізгіштігіне ауысады, демек натрий иондарының жасуша ішіне ағыны артады.
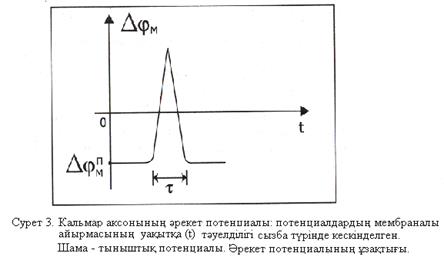
Мембрананың беттері арасындағы потенциалдар айырмасы нөльге дейін көтеріліп, соңынан оң болады – мембрананың деполяризациясы орнайды. Әрі қарай үрдіс кері бағытта жүреді: натрий иондарының жасуша ішіне ағыны біртіндеп кеміп, «қозбаған» мәніне қайтып оралады; сонымен қатар потенциалдар айырмасы да 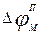 мәніне қайтып оралады. Осындай импульс (серпініс) әрекет потенциалы (сурет 3) деп аталады.
мәніне қайтып оралады. Осындай импульс (серпініс) әрекет потенциалы (сурет 3) деп аталады.
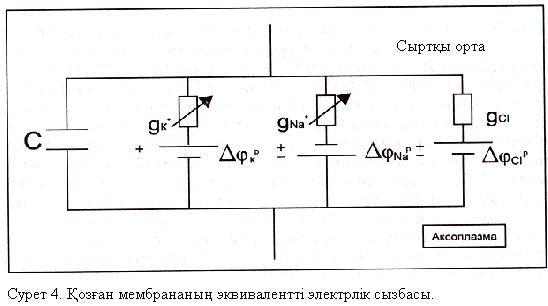 Ходжкин мен Хаксли сонымен бірге қозу үрдісінің электрлік үлгісін жасады, осы үлгіге сәйкес мембрана мынадай сызба түрінде көрсетіледі (сурет 4).
Ходжкин мен Хаксли сонымен бірге қозу үрдісінің электрлік үлгісін жасады, осы үлгіге сәйкес мембрана мынадай сызба түрінде көрсетіледі (сурет 4).
4-ші суретте сыйымдылық конденсаторы С, өткізгіштері 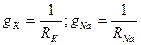 болатын екі айнымалы резистор, Нернст формуласымен есептелінген калий, натрий және хлор иондары үшін тепе- теңдік салмақты потенциалдар айырмасын білдіретін ЭҚК-ң 3 көзі және сәйкес өткізгіштігі сызбада көрсетілген тағы бір кедергі (хлор мен бірқатар басқа иондар үшін мембрананың қосынды кедергісі) көрсетілген.Осы үлгіге сәйкес мембрана арқылы өтетін толық ағын 4 қосылғыштардың қосындысына тең:
болатын екі айнымалы резистор, Нернст формуласымен есептелінген калий, натрий және хлор иондары үшін тепе- теңдік салмақты потенциалдар айырмасын білдіретін ЭҚК-ң 3 көзі және сәйкес өткізгіштігі сызбада көрсетілген тағы бір кедергі (хлор мен бірқатар басқа иондар үшін мембрананың қосынды кедергісі) көрсетілген.Осы үлгіге сәйкес мембрана арқылы өтетін толық ағын 4 қосылғыштардың қосындысына тең: 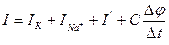 ;
;
мұндағы Iк – калий иондарының ағыны; INa – натрий иондарының ағыны; I’ – хлор иондары мен басқа да иондардың ағыны, оны кейде утечка ағыны деп айтады;
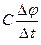 - конденсатор қосылған тізбектің тармағынан ағатын сыйымдылық тогы (мембрараның қайта зарядталу тогы);
- конденсатор қосылған тізбектің тармағынан ағатын сыйымдылық тогы (мембрараның қайта зарядталу тогы);
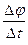 - потенциалдың өзгеру жылдамдығы.
- потенциалдың өзгеру жылдамдығы.
Алдымен калийдің ағынын қарастырайық. Калий иондары мембрана арқылы оның екі жағына әрқашан өтеді. Мембранадағы потенциал ( 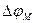 ) басқалай айтқанда калий үшін тепе- теңдік салмақты потенциал деп аталатын
) басқалай айтқанда калий үшін тепе- теңдік салмақты потенциал деп аталатын 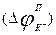 Нернст потенциалына тең болғанда, онда динамикалық тепе-теңдік орнайды, яғни калий ағыны
Нернст потенциалына тең болғанда, онда динамикалық тепе-теңдік орнайды, яғни калий ағыны 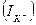 нөльге тең. Егер мембраналы потенциал тепе-теңдік салмақты потенциалдан ауытқыса, онда калий ағыны пайда болып, оның шамасын Ом заңымен анықтауға болады:
нөльге тең. Егер мембраналы потенциал тепе-теңдік салмақты потенциалдан ауытқыса, онда калий ағыны пайда болып, оның шамасын Ом заңымен анықтауға болады: 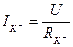 .
.
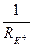 -ды калий өткізгіштігімен
-ды калий өткізгіштігімен 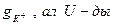 мембраналы потенциалдың тепе-теңдік потенциалдан ауытқу шамасымен, яғни
мембраналы потенциалдың тепе-теңдік потенциалдан ауытқу шамасымен, яғни 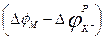 алмастырсақ, онда
алмастырсақ, онда 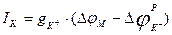 болады.
болады.
Осындай формуламен натрий ағынының күші де анықталады:
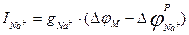 ;
;
мұндағы 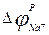 -тепе-теңдік салмақты натрий потенциалы, яғни натрий үшін Нернст потенциалы, оның мәні шамамен +40 мВ-қа тең (ол жуық шамамен овершутқа тең – максимум нүктесінде мәнінің нөльден артуына тең). Ағындар күші мембраналы потенциалдан жеткілікті күрделі тәсілмен байланысады: жақшаның ішінде тұр, сонымен бірге gк мен gNa коэффициенттеріне аргумент түрінде енгізілген.
-тепе-теңдік салмақты натрий потенциалы, яғни натрий үшін Нернст потенциалы, оның мәні шамамен +40 мВ-қа тең (ол жуық шамамен овершутқа тең – максимум нүктесінде мәнінің нөльден артуына тең). Ағындар күші мембраналы потенциалдан жеткілікті күрделі тәсілмен байланысады: жақшаның ішінде тұр, сонымен бірге gк мен gNa коэффициенттеріне аргумент түрінде енгізілген.
Қандайда бір өзгермейтін мембраналы потенциал кезінде gк мен gNa өткізгіштерінің уақытқа тәуелділігін Ходжкин – Хаксли тәжірибелерінде оқып-зерттеген. Өте қысқа уақытта мембранада белгілі бір потенциалдар айырмасын тұрақтандырып, ұзақ уақыт ұстап тұрған. Бұл сыйымдылық токтардан құтылуға мүмкіндік берген: егер 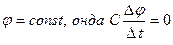 . Калий және натрий иондарын жекелей тіркеу үшін блокаторлар (тосқауылдар) пайдаланған. Тетраэтиламмонийді енгізу кезінде (калийді тосқауылдаушы ) натрий иондары ағынының уақытқа тәуелділігін анықтаған, ал тетродотоксинді енгізу арқылы – калий иондары ағынының уақытқа тәуелділігін анықтаған.
. Калий және натрий иондарын жекелей тіркеу үшін блокаторлар (тосқауылдар) пайдаланған. Тетраэтиламмонийді енгізу кезінде (калийді тосқауылдаушы ) натрий иондары ағынының уақытқа тәуелділігін анықтаған, ал тетродотоксинді енгізу арқылы – калий иондары ағынының уақытқа тәуелділігін анықтаған.
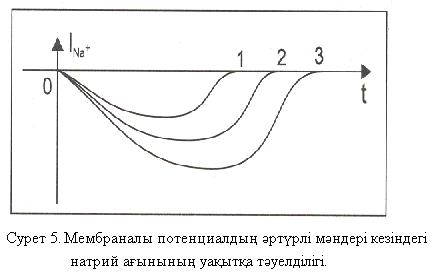
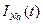 тәуелділігі 5-ші суреттегідей графикті кескінделген. 1, 2 , 3 графиктер анықталған потенциалдардың әртүрлі мәндеріне сәйкес келген. Нақтылық үшін
тәуелділігі 5-ші суреттегідей графикті кескінделген. 1, 2 , 3 графиктер анықталған потенциалдардың әртүрлі мәндеріне сәйкес келген. Нақтылық үшін 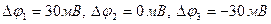 .
.
5-ші сурет біріншіден gNa өткізгіштігі уақытқа байланысты екендігі жөнінде маңызды қорытынды жасауға мүмкіндік береді, себебі потенциалдар айырмасы тұрақты болған кезде 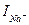 ағыны уақытқа байланысты өзгерген, ал бұл
ағыны уақытқа байланысты өзгерген, ал бұл 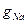 өткізгіштігі уақыттың функциясы болып табылады деген шартта ғана мүмкін :
өткізгіштігі уақыттың функциясы болып табылады деген шартта ғана мүмкін : 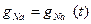 .
.
Екіншіден тепе-теңдік салмақты натрий потенциалының Нернст формуласы бойынша шамасы + 40 мВ болады. Сонда 3-ші жағдай кезінде, потенциалдың анықталған мәні
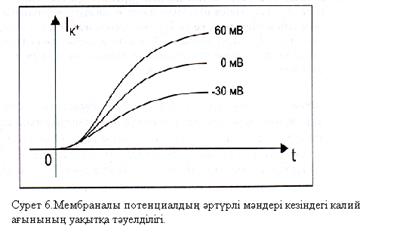
– 30 мВ-қа тең болған, ал 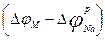 айырмасы өте үлкен (+ 70 мВ) және абсолют шамасы бойынша осы жағдайдағы ағын күші де өте жоғары болады. Бұл Ходжкин – Хаксли үлгісімен үйлесімді.Үшіншіден, өткізгіштігі мембраналы потенциалға байланысты болатындығы туралы қорытынды жасауға болады. Осындай ұқсас қисықтар калий ағыны үшін 6-шы суретте көрсетілгендей түрде болады.
айырмасы өте үлкен (+ 70 мВ) және абсолют шамасы бойынша осы жағдайдағы ағын күші де өте жоғары болады. Бұл Ходжкин – Хаксли үлгісімен үйлесімді.Үшіншіден, өткізгіштігі мембраналы потенциалға байланысты болатындығы туралы қорытынды жасауға болады. Осындай ұқсас қисықтар калий ағыны үшін 6-шы суретте көрсетілгендей түрде болады.
Тәжірибелер әрекет потенциалының бірінші фазасы натрий иондарының жасуша ішіне ағынымен, ал екінші фазасы- калий иондарының жасуша сыртына қарай ағынымен байланысты болатындығын дәлелдеп берді (сурет 7). Одан әрі Ходжкин мен Хаксли өткізгіштіктің өзгеруін түсіндіретін, демек қозу үрдісі кезінде натрий және калий иондарының мембрана арқылы ағынын түсіндіретін математикалық үлгіні ұсынды. Осы үлгінің негізгі постулаттарының бірі: мембранада натрий және калий иондарын тасымалдау үшін жеке арналар болады.
Арналардың негізгі қасиеттері:
- селективтілік (таңдамалалық) - көптеген жағдайда тек бір түрлі иондарды ғана өткізуге қабілеттілігі;
- жеке арналар жұмысының тәуелсіздігі – бір арна арқылы ағын ағының басқа арнамен жүруі, жүрмеуіне байланысты емес;
- иондық арналардың өткізгіштігінің дискреті (үздікті) сипаты: бірінші жуықтауда иондық арна екі күйде болады деп ұйғаруға болады- ашық не жабық, осы күйлер арасындағы алмасулар (өтулер) уақыттың кездейсоқ мезетінде орындалады және статистикалық заңдылықтарға бағынады.
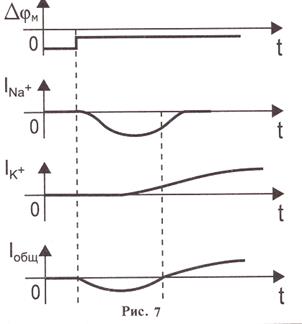 Сурет 7. а-арнайы сызбаның көмегімен мембранада анықталған потенциалдар айырмасының орнатылу; б – тогының уақытқа тәуелділігі. Графиктің теріс аймақта орналасуы натрий иондарының тогы жасуша ішіне бағытталғанын білдіреді. Абсолют шамасы бойынша ол алдымен артып, одан кейін кемиді; в –
Сурет 7. а-арнайы сызбаның көмегімен мембранада анықталған потенциалдар айырмасының орнатылу; б – тогының уақытқа тәуелділігі. Графиктің теріс аймақта орналасуы натрий иондарының тогы жасуша ішіне бағытталғанын білдіреді. Абсолют шамасы бойынша ол алдымен артып, одан кейін кемиді; в – 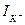 тогының уақытқа тәуелділігі. Қисық сызығының оң аймақта орналасуы калий иондары тогының жасуша сыртына бағытталғандығын білдіреді.
тогының уақытқа тәуелділігі. Қисық сызығының оң аймақта орналасуы калий иондары тогының жасуша сыртына бағытталғандығын білдіреді.
-ң артуы анықталған потенциалдар айырмасы орнатылғаннан біршама кейін, инактивтелген кезде басталады, ; г – мембрана арқылы 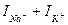 қосынды тогы. Қисықтың абсцисса осін қиюына сәйкес келетін уақыт мезеті кезінде ол 0-ге тең, содан кейін бағытын өзгертеді.
қосынды тогы. Қисықтың абсцисса осін қиюына сәйкес келетін уақыт мезеті кезінде ол 0-ге тең, содан кейін бағытын өзгертеді.
Ток күшінің әрбір жеке арна арқылы секірмелі өзгеретініне қарамастан, қосынды токтың уақытқа тәуелділігі баяу қисық болады, бұл бір мезгілде қызмет жасайтын арналардың көп санымен анықталады (мысалы, ұзындығы 1 см және диаметрі 1 мм болатын кальмар аксонында шамамен 1010 арна бар). 8-ші суретте жеке натрий арналарындағы токтар мен қосынды натрий тогының арасындағы байланыс көрсетілген- арна параметрлерінің мембраналы потенциалға тәуелділігі. Бұл мембрананың деполяризациясы басталғаннан кейін сәйкес токтар бір не басқа кинетика бойынша өзгере бастайтындығынан білінеді. Иондық арналар туралы гипотеза бойынша осы үрдіс келесі түрде жүзеге асады. Ионды селективті арнаның сенсор деп аталатын құрылымды элементі болады. Осы элемент электр өрісінің әсеріне сезімтал. Мембраналы потенциал өзгерген кезде, оған әсер ететін күштің шамасы өзгереді, нәтижесінде иондық арнаның осы бөлігі орын ауыстырып, қақпаның (бәрі не ешқайсысы емес заңымен жүретін өзіндік бір қалқалар) ашылып-жабылу ықтималдығын өзгертеді. Шамамен 1952ж. Хаксли жасалған математикалық үлгі бойынша мембраналы потенциалдар айырмасының уақытқа тәуелді қалай өзгертіндігіне есептеген. Осы есептеудің нәтижесі тәжірибе жүзінде тіркелген әрекет потенциалымен жоғары дәлдікпен сәйкес келді.
Ходжкин – Хаксли үлгісі рефрактерлік (қозу кезінде қозушылықтың өзгеруі), қозу табалдырығы және т.б. құбылыстарды ашты және тәжірибеде бақыланған әрекет потенциалының қасиеттерін теориялық жолмен түсіндіруге мүмкіндік берді:
- деполяризацияланатын потенциалдың шекті (табалдырықты) мәні;
- «барлығы не ешқайсысы емес»: егер деполяризацияланатын потенциал шекті мәнінен үлкен болса, онда әрекет потенциалы дамиды, оның амплитудасы қоздырушы импульстің амплитудасына байланысты болмайды; егер деполяризацияланатын потенциалдың амплитудасы шекті мәнінен кіші болса, онда әрекет потенциалы пайда болмайды ;
- әрекет потенциалының дамуы кезінде және қозудың тоқтауынан кейінгі құбылыстар кезінде мембрананың рефрактерлік периодының болуы (қозбаушылық ). Жоғарыда айтылғандай, тынышытық потенциалы жасушаның іші мен сыртындағы калий иондарының мөлшерінің айырмашылығы нәтижесінде пайда болады. Бірақ бірінші фазада натрий иондары ағынының жасуша ішіне және екінші фаза кезінде жасуша сыртына қарай калий иондары ағынының сүйемелдеуімен жүретін бірнеше әрекет потенциалдарынан кейін, жасушаның іші мен сыртындағы осы иондардың мөлшерінің айырмашылығы да өзгеру керек. Осыған байланысты, мысалы, Нернст формуласындағы логарифм таңбасы астындағы калий иондарның ішкі және сыртқы концентрацияларының қатынасының кемуі нәтижесінде, тыныштық потенциалының шамасы біртіндеп 0-ге жақындауы керек. Бірақ бұл орындалмайды. Өйткені концентрация айырмашылығын ұстап тұратын қандайда бір механизмнің болуы мүмкін. Осындай механизмнің көмегімен калий иондары жасуша ішіне өтулері керек (жасушада калий иондарының онсыз да мөлшері көп), ал натрий иондары жасуша сыртына шығарылуы керек, ондағы олардың мөлшері ішіндегіден онсыз да көп. Бұл қандай механизм?
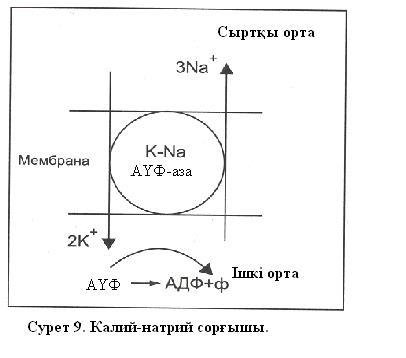
Жасушалы мембранада әртүрлі ақуыз молекулалары бар. Олардың кейбіреулері өзінше бір сорғыштың (насос) ролін атқарады екен, олар калий иондарын жасуша ішіне енгізеді (сорады), ал натрий иондарын жасуша сыртына сорып шығарады (белсенді ионды тасымалдың мысалы). Сорғыштардың қандай көздің энергиясы есебінен жұмыс істейтіндігі белгілі. Калий – натрий сорғышы жағдайында радиоактивті изотоптармен тәжірибелер жасуша ішіне калийдің 2 ионын енгізу үшін, ал сыртқа 3 натрий ионын сорып шығару үшін (сорғыш жұмысының бір циклі ) АҮФ-ң бір молекуласының ыдырау энергиясы жеткілікті екендігін көрсетті (сурет 9). Бұны 1957ж . С.Скоу байқаған болатын.
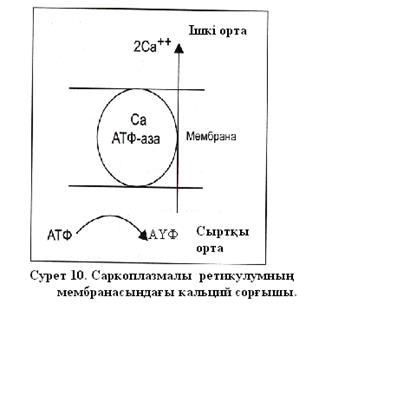
Иондық сорғыштардың жұмысы жасушаның іші мен сыртындағы иондар концентрациясымен басқарылатындығы анықталды. Сорғыш жұмысы жасуша сыртындағы калий иондарының көпболуымен және ішіндегі натрий иондарының артық болумен үдетіледі. Жасушаның өртүрлі қызметтерінің жүзеге асырылуында кальций ионы маңызды роль атқарады. Бұлшыеттің жиырылуы үшін көп мөлшерде кальций иондары қажет және оны жасуша денесін жарып орналасқан әрбір ақуыз фибрильдеріне жеткізу керек, содан кейін бұлшықет босаңсу үшін оны фибрильдерден жылдам алу керек. Егер әрбір кальций ионы жасушаның сыртқы мембранасы арқылы келіп және кететін болса, онда оның жылдам орын ауыстыру мүмкін болмас еді. Бұлшықеттің жасушаларының ішінде арнайы ішкі мембранадан құралған тармақты қуыс пен түтікшелер жүйесі болады, оларда кальций сақталады және бұлшықет босаңсығанда кальций иондары қайтадан оларға келеді. Барлық осы мембрана кальций сорғыштармен «қапталған». Кальцийдің белсенді тасымалының көзі сонымен бірге АҮФ гидролизі болып табылады ( сурет10). Жүйке талшықтарымен әрекет потенциалының таралуын қарастырайық. Егер қозған мембрананың қандайда бір бөлігінде әрекет потенциалы қалыптасса , мембрана деполяризацияланады, қозу мембрананың басқа да бөліктеріне тарайды.
Жүйке серпінісінің аксонмен берілуінің таралуын қарастырайық. Аксоплазмада және қоршаған ерітіндіде мембрананың бетіндегі потенциалы жоғары бөліктер (оң зарядталған) мен потенциалы кіші (теріс зарядталған) бөліктер аралығында локальді (жергілікті) токтар пайда болады.Тірі ұлпаларда қозудың нәтижесінде пайда болатын токтар локальді деп аталады, себебі олар қозған бөліктен біраз ғана аралыққа таралады. Локальді токтар аксонның ішінде де, оның сыртқы бетінде де пайда болады. Локальді электрлік токтар мембрананың қозбаған бөлігінің ішкі бетіндегі потенциалдың 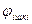 артуына және қозған бөлікпен көршілес болатын мембрананың қозбаған бөлігінің сыртқы потенциалының
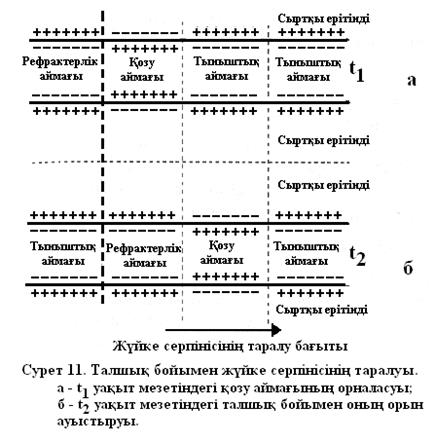
артуына және қозған бөлікпен көршілес болатын мембрананың қозбаған бөлігінің сыртқы потенциалының 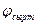 төмендеуіне әкеліп соғады. Сөйтіп, теріс тыныштық потенциалы абсолют шамасы бойынша кемиді, яғни шекті мәнінен жоғары артады. Сондықтан мембараналы потенциалдың өзгеруі әсерінен натрий арналары ашылып, оның әрі қарай артуы мембрана арқылы иондар ағынының нәтижесінде жүзеге асады. Мембрана деполяризацияланып, әрекет потенциалы дамиды. Одан соң қозу алғашқы мембрананың алғашқы қозбаған бөліктеріне беріледі (сурет11).
төмендеуіне әкеліп соғады. Сөйтіп, теріс тыныштық потенциалы абсолют шамасы бойынша кемиді, яғни шекті мәнінен жоғары артады. Сондықтан мембараналы потенциалдың өзгеруі әсерінен натрий арналары ашылып, оның әрі қарай артуы мембрана арқылы иондар ағынының нәтижесінде жүзеге асады. Мембрана деполяризацияланып, әрекет потенциалы дамиды. Одан соң қозу алғашқы мембрананың алғашқы қозбаған бөліктеріне беріледі (сурет11).
Кальмар аксонымен жүйке серпінісінің таралу жылдамдығының үлкен болуын омыртқалылар аксондарымен салыстырғандағы үлкен диаметрі қамтамасыз етеді. Омыртқалыларда жүйке талшықтарымен қозудың үлкен таралу (берілу) жылдамдығы басқа тәсілмен қамтамасыз етіледі. Омыртқалылар аксондары миелинді қабықшамен жабдықталған, ол мембрананың кедергісін арттырады. Миелиндірілген талшық арқылы қозу Рантьенің бір қармауынан ( миелин қабықшасы жоқ бөліктен) екіншісіне сальтаторлы (секірмелі) таралады. Жүйке серпіністері аксондармен қандай да бір дәрежеде электр сигналдарының кабельді- релейлі сымдармен таралуына ұқсас таралады. Электр серпінісі аралық релейлі станцияларда осы серпіністің күшеюі есебінен, өшпей беріледі, ал осы станциялардың ролін аксондарда әрекет потенциалы өндірілетін қозған мембрананың бөліктері атқарады.
Дата добавления: 2015-05-26; просмотров: 3752;