Загальна схема
Першою моделлю потоків енергії в екосостемі була схема трофічних шляхів для прерії (В.Шелфолд, 1913). Кількісно характеристику потоку енергії по всій трофічній структурі вперше виконав Р.Ліндеман (1942).Згодом подібні дослідження для біотичних елементів виконали Г.Одум (1957,1960), А.Макфедьєн (1963), П.Дювіньйо (1964), а для абіотичних дослідження радіаційного балансу різних типів ландшафтів проводили М.І.Будико, Ю.Р.Раунер і ін.
Основним джерелом енергії для багатьох процесів у геосистемі є сонячна. Порівняно з нею енергії до геосистеми від інших джерел надходить дуже мало (теплової енергії з надр Землі – 0,04% сумарної сонячної радіації, тектонічних рухів – 0,0005%). При цьому сонячна енергія і використовується в геосистемі найбільш ефективно: вона здатна трансформуватися в інші види енергії (теплову, хімічну, механічну), завдяки їй відбувається продукування біомаси, вологообмін, циркуляція повітряних мас тощо.
Загальну схему потоку і трансформації сонячної енергії в геосистемі наведено на рис.7. На верхню межу атмосфери надходить 2 кал/см2 ·хв сонячної енергії. Проходячи крізь атмосферу, вона послаблюється атмосферними газами та пилом. При цьому ступінь послаблення залежить від довжини хвилі (частоти) світла. З екологічної точки зору, найбільш важливими обставинами диференційованого послаблення випромінювання є дві: 1) ультрафіолетове випромінювання (найбільш небезпечне для протоплазми) практично не проходить крізь озоновий шар, що й забезпечує можливість життя на планеті; 2) менше всього послаблюється видиме світло, що необхідне для фотосинтезу, а тому він може відбуватись і в похмурі дні.
Сумарна радіація (R на рис.7) складається з прямої (RI) та розсіяної (RS). Співвідношення між ними залежить від географічного положення геосистеми, хмарності та інших кліматичних факторів. Це співвідношення визначає ряд похідних ландшафтно-екологічних факторів, зокрема – експозиційний. Чим більша частка прямої радіації в сумарній, тим більше значення має фактор експозиції схилу.
Частина сумарної радіації, досягаючи геосистеми, витрачається на ефективне випромінювання в атмосферу RE та відбивається поверхнею геосистеми (відбита радіація RA). Величина останнього потоку визначається характером поверхні геосистеми (її альбедо) і суттєво змінюється в різних умовах: у дібровах – 0,14 – 0,17, степах – 0,20 – 0,23, на солончаках – 0,35, засніженій поверхні – до 0,95.
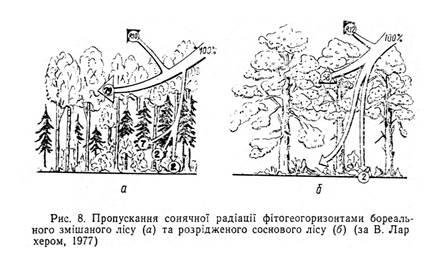
Альбедо слід розглядати як інтегральний параметр вертикальної структури геосистеми, який визначає потік відбитої радіації з неї як з деякого нерозчленованого цілого. Насправді ж потік сонячної радіації, проходячи від верхньої межі геосистеми до поверхні грунту, значно змінює свою інтенсивність, спектральний склад та інші характеристики. Ці зміни визначаються геогоризонтальною вертикальною структурою геосистеми,
особливо потужністю та часткою геомас листя в окремих аерофітогоризонтах (рис.8).
Ефективне випромінювання та відбита радіація втрачаються для геосистеми, а та частина сумарної радіації, що безпосередньо йде на різні процеси в геосистемі, називається її радіаційним балансом RB. Більша його частина витрачається на випаровування (фізичне Е і трансформацію Т) та на

 турбулентну віддачу атмосфері Q, тобто на забезпечення вологообміну та прогрівання повітря геосистеми.
турбулентну віддачу атмосфері Q, тобто на забезпечення вологообміну та прогрівання повітря геосистеми.
Витрати тепла на фотосинтез Р становлять дуже малу частку радіаційного балансу – в середньому 1,3%. Проте її роль у геосистемі надзвичайно велика, бо власне вона зумовлює продуційний та інші важливі біотичні процеси. При фотосинтезі використовується фотосинтетично активна радіація ФАР, частка якої становить близько 45% сумарної (40% прямої та 62% розсіяної). Рослинний покрив поглинає 90% світлової енергії ФАР, проте переважна його частина йде на транспірацію та регулювання температури рослин і лише 0,5 – 1,5 % на фотосинтез. Ефективність фотосинтезу визначається гідротермічними умовами геосистем. Найбільша вона за максимальної теплозабезпеченості при оптимальному співвідношенні тепла й вологи. Такі природні умови характерні для екваторіальних лісів, де ефективність фотосинтезу досягає 4,5% ФАР, або 1,5% сумарної радіації. У штучних умовах можна довести ефективність фотосинтезу до максимальної – 34% ФАР.
Енергія Р, що пішла на забезпечення реакції фотосинтезу, витрачається на дихання рослин Рb (близько 50%), а решта становить чисту первинну продукцію Ра - накопичення енергії в рослинній біомасі. З цієї енергії деяка її частина Рm з відмиранням рослин або їх окремих органів переходить до мортмаси, частина Рі залишається у фітомасі й зумовлює її щорічний приріст, частина Z разом з фітомасою, що поїдається тваринами, переходить до наступного трофічного рівня. Співвідношення мі цими статтями енергетичних витрат залежить від багатьох факторів. З них особливо важливі видовий склад рослинного угруповання, вікова структура популяцій, їх екологічний стан, характер господарського використання, чисельність та популяційний склад тваринного населення біоценозу.
Енергія Z, що надходить з фітомасою до травоїдних тварин, частково йде на потреби самих цих тварин, а частково переходить до тварин-хижахів. Ці тварини, в свою чергу, поїдаються хижаками вищих порядків, а відтак передають частину енергії і їм. Так формується трофічна структура геосистеми. В екології при її дослідженні прийнято виділяти трофічні рівні – сукупність організмів, які отримують енергію від рослин через однакове число організмів-посередників. Самі зелені рослини формують перший трофічний рівень – рівень продуцентів (або автотрофів); травоїдні тварини (зайці, олені тощо) – другий рівень (первинних консументів, або гетеротрофів 1-го порядку); хижаки – третій (рівень вторинних консументів, або гетеротрофів 3-го порядку) і так далі. Оскільки потік енергії від одного трофічного рівня до вищого супроводжується її втратами (до 95%), кількість трофічних рівнів не може бути необмеженою і лише в енергетично багатих екосистемах може досягати 5-6 (екваторіальні ліси, морські планктонові екосистеми).
Потік енергії Z від одного організму до іншого супроводжується її втратами. Їжа як рослинного, так і тваринного походження повністю організмом не засвоюється. Частина її виводиться з організму у вигляді екскрементів Ze, частина Zb витрачається на дихання тварин, а решта енергії Zi може йти на синтез нової біомаси (точніше – зоомаси) в результаті росту організму та розмноження. З цієї енергії Zm втрачається популяцією внаслідок загибелі її особин і далі включається в енергетичні потоки переробки мортмаси, а решта енергії Zz переходить до організмів –консументів вищих рівнів, де структура енергетичних потоків аналогічна. Величина потоку Z залежить насамперед від чисельності тварин та типу біому. Взагалі у трав’яних геосистемах відсоток наземної рослинної продукції, що споживається травоїдними тваринами, більший, ніж у лісових. У хвойних він становить 2-3%, широколистих – до 3, в степових геосистемах досягає 8-10, в саванах Африки 28-60, а на культурних пасовищах США 30-45%.
Потоки та трансформація енергії, що міститься у відмерлих частинах рослин Рm, екскрементах консументів Ze та їх трупах Zm, ще не з’ясовані до кінця. Сукупність цих потоків прийнято називати детритним циклом геосистеми.
Деяка частина енергії, що міститься у відмерлій фітомасі, втрачається через її окислення (переважно фотохімічне), в результаті чого виділяється СО2. У лучному степу Курського заповідника, наприклад, ця частка становить 4% енергії відмерлої надземної фітомаси. У лісових геосистемах спостерігаються втрати енергії (10-30%), зумовлені вимиванням дощовою водою органічних речовин із свіжовпалого листя. У схилових геосистемах значна частка енергії Рm втрачається з виносом її за межі геосистеми горизонтальними потоками води. Проте більшу роль у трансформації енергії в детритному циклі відіграють не абіотичні фактори, а сапрофаги (синоніми – редуценти, детритні консументи) – живі організми, що живляться неживим органічним матеріалом, який можна метаболізувати для отримання енергії. У наземних геосистемах сапрофаги споживають до 90-95% усієї чистої первинної продукції. До них належать такі великі тварини, як грифи, ворони, краби, а також черви, кліщі, бактерії та гриби.
Більша частина енергії в детритному циклі витрачається сапротрофами на дихання, а решта переходить спочатку в проміжні продукти розкладу органічної речовини та неспецифічну органічну речовину грунту. У лучному степу в них залишається не більше 50% енергії, що надходить з рослинними рештками і екскрементами. На наступному етапі трансформації цієї енергії до 70% її втрачається на мінералізацію органічної речовини мікроорганізмами та до 25% на їх дихання. Решта переходить у специфічні гумусові речовини – продукти гуміфікації. Таким чином, у гумус переходить не більше 5-10% енергії рослинних решток.
Дата добавления: 2015-05-16; просмотров: 2245;