ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
А. Структурно-функциональная организация коры. Кора большого мозга представляет собой многослойную нейронную ткань с множеством складок. В коре выделяют 6 слоев (I—VI), каждый из которых состоит из пирамидных и звездчатых клеток
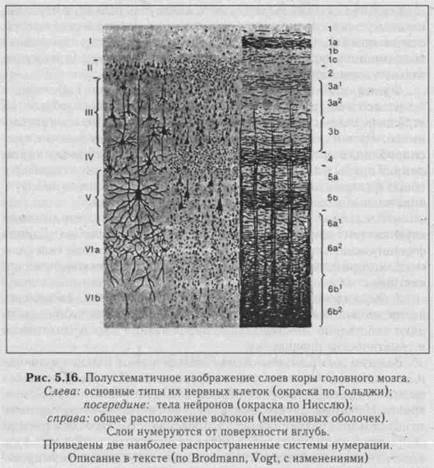
(рис. 5.16). Главная особенность пирамидных клеток (название отражает форму клеток) заключается в том, что их аксоны выходят из коры, а также оканчиваются в других корковых структурах. Название звездчатых клеток также обусловлено их формой; их аксоны оканчиваются в коре, т. е. речь идет о корковых интернейронах. ВI—IV слоях происходят восприятие и обработка поступающих в кору сигналов в виде импульсов. Покидающие кору эфферентные пути формируются преимущественно в У-У1 слоях. Деление коры на различные поля проведено К. Бродманом (1909) на основе цито-архитектонических признаков - формы и расположения нейронов. Автор выделил 52 поля, многие из них характеризуются функциональными и нейрохимическими особенностями.
Кортикализация функций — возрастание в филогенезе роли коры большого мозга в анализе и регуляция функций организма и подчинение себе нижележащих отделов ЦНС. Выключение коры больших полушарий у обезьян и у человека приводит к потере не только локомоции, но и выпрямительных рефлексов, которые у других млекопитающих сохраняются.
Б. Локализация функций в коре большого мозга. В определенных участках коры большого мозга сосредоточены нейроны, воспринимающие определенный вид раздражителя: свет - затылочная область, звук - височная доля и т.д. Двигательные функции также распределены на значительных площадях коры большого мозга, например, нейроны, аксоны которых формируют пирамидный тракт, расположены не только в моторных областях, но и за их пределами. В настоящее время принято подразделение коры на сенсорные, двигательные и ассоциативные (неспецифические) зоны.
Сенсорные зоны коры — это зоны, в которые поступает сенсорная информация (от рецепторов): проекционная кора, корковые отделы анализаторов по И. П. Павлову. Эти зоны расположены преимущественно в теменной, височной и затылочной долях. Восходящие пути в сенсорную кору поступают преимущественно от релейных сенсорных ядер таламуса. Первичные сенсорные области - зоны сенсорной коры, раздражение или разрушение которых вызывает четкие и постоянные изменения чувствительности организма (ядра анализаторов по И. П. Павлову). Они состоят преимущественно из мономодальных нейронов и формируют ощущения одного качества (одной модальности). В первичных сенсорных зонах обычно имеется четкое пространственное (топографическое) представительство частей тела, их рецепторных полей. Вокруг первичных сенсорных зон находятся менее локализованные вторичные сенсорные зоны, нейроны которых отвечают на действие нескольких раздражителей, т.е. они полимодальны.
Важнейшей сенсорной областью являются теменная кора постцентральной извилины и соответствующая ей часть парацент-ральной дольки на медиальной поверхности полушарий, которая обозначается как соматосенсорная область I. Здесь имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептив-ной чувствительности и чувствительности опорнодвигательного аппарата - от мышечных, суставных, сухожильных рецепторов (рис. 5.17).
Кроме соматосенсорной области I выделяют меньших размеров соматосенсорную область II, расположенную на границе

|

|
Двигательные зоны коры. Выделяют первичную и вторичную моторную области. В первичной моторной коре (прецентраль-ная извилина, поле 4) расположены нейроны, иннервирующие мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела (см. рис. 5.17). Основной закономерностью топографического представительства является то, что регуляция деятельности мышц, обеспечивающих наиболее точные и разнообразные движения (речь, письмо, мимика), требует участия больших по площади участков двигательной коры.
Вторичная двигательная кора (поле 6) расположена как на латеральной поверхности полушарий, впереди прецентральной извилины (премоторная кора), так и на медиальной поверхности, соответствующей коре верхней лобной извилины (дополнительная моторная область). Вторичная двигательная кора в функциональном плане имеет главенствующее значение по отношению к первичной двигательной коре, осуществляя высшие двигательные функции, связанные с планированием и координацией произвольных движений. Здесь в наибольшей степени регистрируется медленно нарастающий отрицательный потенциал готовности, возникающий примерно за 1 с до начала движения. Кора поля 6 получает основную часть им-пульсации от базальных ганглиев и мозжечка, участвует в переко- " дировании информации о плане сложных движений.
Ассоциативные области коры (синонимы: неспецифическая, межсенсорная, межанализаторная кора) включают участки новой коры большого мозга, которые расположены вокруг проекционных зон и рядом с двигательными зонами, но не выполняют непосредственно чувствительных или двигательных функций - им нельзя приписывать преимущественно сенсорных или двигательных функций, нейроны этих зон обладают большими способностями к обучению. Границы этих областей обозначены недостаточно четко. Нейроны ассоциативной коры являются полисенсорными (полимодальными): они отвечают, как правило, не на один (как нейроны первичных зон), а на несколько раздражителей - один и тот же нейрон может возбуждаться при раздражении слуховых, зрительных, кожных и других рецепторов. В результате этого ассоциативная кора представляет собой своеобразный коллектор различных сенсорных возбуждений, участвует в интеграции сенсорной информации и в обеспечении взаимодействия сенсорных и моторных областей коры. В настоящее время по таламо-кортикаль-ным проекциям выделяют две основные ассоциативные системы мозга: таламо-теменную и таламо-лобную. Существует мнение о целесообразности выделения и третьей - таламо-височной системы.
Раздел II
ФИЗИОЛОГИЯ ВНУТРЕННИХ ОРГАНОВ И СИСТЕМ ОРГАНИЗМА
Дата добавления: 2015-05-16; просмотров: 911;