Генетическое разнообразие
Естественное богатство нашей планеты связано с разнообразием генетических вариаций. Генетическое разнообразие, т. е. поддержание генотипических гетерозиготности, полиморфизма и другой генотипической изменчивости, которая вызвана адаптационной необходимостью в природных популяциях, представлено наследуемым разнообразием внутри и между популяциями организмов.
Как известно, генетическое разнообразие определяется варьированием последовательностей 4 комплиментарных нуклеотидов в нуклеиновых кислотах, составляющих генетический код. Каждый вид несет в себе огромное количество генетической информации: ДНК бактерии содержит около 1 000 генов, грибы – до 10 000, высшие растения – до 400 000. Огромно количество генов у многих цветковых растений и высших таксонов животных. Например, ДНК человека содержит более 30 тыс. генов.
Новые генетические вариации возникают у особей через генные и хромосомные мутации, а также у организмов, которым свойственно половое размножение, через рекомбинацию генов. Генетические вариации могут быть оценены у любых организмов, от растений до человека, как число возможных комбинаций различных форм от каждой генной последовательности. Другие разновидности генетического разнообразия, например количество ДНК на клетку, структура и число хромосом, могут быть определены на всех уровнях организации живого.
Огромное множество генетических вариаций представлено у скрещивающихся популяций и может быть осуществлено посредством селекции. Различная жизнеспособность отражается в изменениях частот генов в генофонде и является реальным отражением эволюции. Значение генетических вариаций очевидно: они дают возможность осуществления и эволюционных изменений и, если это необходимо, искусственного отбора.
Только небольшая часть (около 1%) генетического материала высших организмов изучена в достаточной мере, когда мы можем знать, какие гены отвечают за определенные проявления фенотипа организмов. Для большей части ДНК ее значение для вариации жизненных форм остается неизвестным.
Каждый из 109 различных генов, распределенных в мировой биоте, не дает идентичного вклада в формирование разнообразия. В частности, гены, контролирующие фундаментальные биохимические процессы, являются строго консервативными у различных таксонов и, в основном, демонстрируют слабую вариабельность, которая сильно связана с жизнеспособностью организмов.
Если судить об утере генофонда с точки зрения генной инженерии, принимая во внимание то, что каждая форма жизни уникальна, вымирание всего лишь одного дикого вида означает безвозвратную потерю от тысячи до сотен тысяч генов с неизвестными потенциальными свойствами. Генная инженерия могла бы использовать это разнообразие для развития медицины и создания новых пищевых ресурсов. Однако разрушение местообитаний и ограничение размножения многих видов приводит к опасному уменьшению генетической изменчивости, сокращая их способности адаптироваться к загрязнению, изменениям климата, болезням и другим неблагоприятным факторам. Основной резервуар генетических ресурсов – природные экосистемы – оказался значительно измененным или разрушенным. Уменьшение генотипического разнообразия, происходящее под воздействием человека, ставит на грань риска возможность будущих адаптаций в экосистемах.
Изучение закономерностей распределения генотипов в популяциях начато Пирсоном [1904]. Он показал, что при наличии разных аллелей одного гена и действия свободного скрещивания в популяциях возникает совершенно определенное распределение генотипов, которое можно представить в виде:
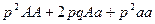 ,
,
где p – концентрация гена A; q – концентрация гена a.
Х. Харди (1908) и В. Вайнберг (1908), специально исследовав это распределение, высказали мнение, что оно является равновесным, так как при отсутствии факторов, нарушающих его, оно может сохраняться в популяциях неограниченное время. Так стала развиваться популяционная генетика. Главная заслуга в разработке популяционной генетики, а особенно ее теоретического и математического аспектов, в этот ранний период [1920–1940 гг.] принадлежит С. С. Четверикову, С. Райту, Р. Фишеру, Дж. Холдейну, А. С. Серебровскому и Н. П. Дубинину.
Биологическая эволюция – это процесс накопления изменений в организмах и увеличение их разнообразия во времени. Эволюционные изменения затрагивают все стороны существования живых организмов: их морфологию, физиологию, поведение и экологию. В основе всех этих изменений лежат генетические изменения, т.е. изменения наследственного вещества, которое, взаимодействуя со средой, определяет все признаки организмов. На генетическом уровне эволюция представляет собой накопление изменений в генетической структуре популяций.
Эволюцию на генетическом уровне можно рассматривать как двухступенчатый процесс. С одной стороны, возникают мутации и рекомбинации – процессы, обусловливающие генетическую изменчивость; с другой стороны, наблюдается дрейф генов и естественный отбор – процессы, посредством которых генетическая изменчивость передается из поколения в поколение.
Эволюция возможна только в том случае, если существует наследственная изменчивость. Единственным поставщиком новых генетических вариантов служит мутационный процесс, однако эти варианты могут по-новому рекомбинироваться в процессе полового размножения, т. е. при независимом расхождении хромосом и вследствие кроссинговера. Генетические варианты, возникшие в результате мутационного и рекомбинационного процессов, передаются из поколения в поколение отнюдь не с равным успехом: частота некоторых из них может увеличиваться за счет других. Помимо мутаций к процессам, изменяющим частоты аллелей в популяции, относится естественный отбор, поток генов (т. е. миграции их) между популяциями и случайный дрейф генов.
На первый взгляд может показаться, что особи с доминантным фенотипом должны встречаться чаще, чем с рецессивным. Однако соотношение 3:1 соблюдается лишь в потомстве двух особей, гетерозиготных по одним и тем же двум аллелям. При других типах скрещивания в потомстве происходит иное расщепление признаков, и такие скрещивания также влияют на частоты генотипов в популяции. Законы Менделя ничего не говорят нам о частотах генотипов в популяциях. Именно об этих частотах идет речь в законе Харди – Вайнберга. Основное утверждение закона Харди – Вайнберга состоит в том, что в отсутствие элементарных эволюционных процессов, а именно мутаций, отбора, миграции и дрейфа генов, частоты генов остаются неизменными из поколения в поколение. Этот закон утверждает также: если скрещивание случайно, то частоты генотипов связаны с частотами генов простыми (квадратичными) соотношениями. Из закона Харди – Вайнберга вытекает следующий вывод: если частоты аллелей у самцов и самок исходно одинаковы, то при случайном скрещивании равновесные частоты генотипов в любом локусе достигаются за одно поколение. Если частоты аллелей у двух полов исходно различны, то для аутосомных локусов они становятся одинаковыми в следующем поколении, поскольку и самцы, и самки получают половину своих генов от отца и половину – от матери. Таким образом, равновесные частоты генотипов достигаются в этом случае за два поколения. Однако в случае сцепленных с полом локусов равновесные частоты достигаются лишь постепенно.
Закон Харди – Вайнберга сформулировали в 1908 году независимо друг от друга математик Г. Х. Харди в Англии и врач В. Вайнберг в Германии. Чтобы понять смысл этого закона, можно привести следующий простой пример. Предположим, что данный локус содержит один из двух аллелей, A и a, представленных с одинаковыми для самцов и самок частотами: p для A и q для a. Представим себе, что самцы и самки скрещиваются случайнымобразом, или, что то же самое, гаметы самцов и самок образуют зиготы, встречаясь случайно. Тогда частота любого генотипа будет равна произведению частот соответствующих аллелей. Вероятность того, что некоторая определенная особь обладает генотипом AA, равна вероятности (p) получить аллель A от матери, умноженной на вероятность (p) получить аллель A от отца, т. е. 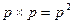 .
.
Закон Харди – Вайнберга гласит, что процесс наследования преемственности сам по себе не ведет к изменению частот аллелей и (при случайном скрещивании) частот генотипов по определенному локусу. Более того, при случайном скрещивании равновесные частоты генотипов по данному локусу достигаются за одно поколение, если исходные частоты аллелей одинаковы у обоих полов.
Равновесные частоты генотипов задаются произведениями частот соответствующих аллелей. Если имеются только два аллеля, A и a, с частотами p и q, то частоты всех трех возможных генотипов выражаются уравнением:
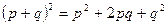
|
| A a AA Aa aa, |
где буквам во второй строке, обозначающим аллели и генотипы, соответствуют расположенные над ними частоты в первой строке.
Если имеется три аллеля, скажем, A1, A2 и A3 , с частотами p, q и r, то частоты генотипов определяются следующим образом:
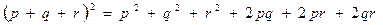
|
| A1 A2 A3 A1A1 A2A2 A3A3 A1A2, A1A3 A2A3. |
Аналогичный прием возведения в квадрат многочлена может быть использован для определения равновесных частот генотипов при любом числе аллелей. Заметим, что сумма всех частот аллелей так же как и сумма всех частот генотипов, должна быть равна единице. Если имеются только два аллеля с частотами p и q, то p + q = 1, и, следовательно, 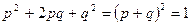 ; если же имеются три аллеля с частотами p, q и r, то p + q + r = 1, и, следовательно, также
; если же имеются три аллеля с частотами p, q и r, то p + q + r = 1, и, следовательно, также 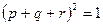 и т. д.
и т. д.
Организмы, обладающие удачными вариантами признаков, имеют большую вероятность по сравнению с другими организмами выжить и оставить потомство. Вследствие этого полезные вариации в ряду поколений будут накапливаться, а вредные или менее полезные вытесняться, элиминироваться. Это и называется процессом естественного отбора, который играет ведущую роль в определении направления и скорости эволюции.
Прямая взаимосвязь между степенью генетической изменчивости в популяции и скоростью эволюции под действием естественного отбора была доказана математическим путем Р. Фишером [1930] в его фундаментальной теореме естественного отбора. Фишер ввел понятие приспособленности и доказал, что скорость возрастания приспособленности популяции в любой момент времени равна генетической вариансе приспособленности в тот же момент времени. Однако прямые доказательства этого факта были получены лишь в конце 60-х годов ХХ столетия.
Мутационный процесс служит источником появления новых мутантных аллелей и перестроек генетического материала. Однако возрастание их частоты в популяции под действием мутационного давления происходит крайне медленно, даже в эволюционном масштабе. К тому же подавляющее большинство возникающих мутаций устраняются из популяции в течение немногих поколений уже в силу случайных причин. Неизбежность такого течения событий впервые обосновал Р. Фишер в 1930 году.
Для человека и других многоклеточных показано, что мутации обычно возникают с частотой от 1 на 100 000 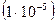 до 1 на 1 000 000
до 1 на 1 000 000 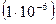 гамет.
гамет.
Новые мутанты, хотя и довольно редко, но постоянно появляются в природе, поскольку существует множество особей каждого вида и множество локусов в генотипе любого организма. Например, число особей того или иного вида насекомых обычно составляет около 100 млн. (108). Если предположить, что средняя мутабельность по одному локусу равна 1 мутации на 100 000 (10-5) гамет, то среднее число вновь возникающих в каждом поколении мутантов по этому локусу для данного вида насекомых составит 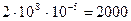 . (Частота возникновения мутаций умножается на число особей и еще на два, так как любая особь представляет собой продукт слияния двух гамет.) В генотипе человека имеется около 100 000 (105) локусов. Предположим, что у человека темп мутирования такой же, как у дрозофилы; в этом случае вероятность того, что генотип каждого человека содержит новый аллель, отсутствовавший в генотипе его родителей, равна
. (Частота возникновения мутаций умножается на число особей и еще на два, так как любая особь представляет собой продукт слияния двух гамет.) В генотипе человека имеется около 100 000 (105) локусов. Предположим, что у человека темп мутирования такой же, как у дрозофилы; в этом случае вероятность того, что генотип каждого человека содержит новый аллель, отсутствовавший в генотипе его родителей, равна 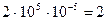 . Иными словами, каждый человек в среднем несет около двух новых мутаций.
. Иными словами, каждый человек в среднем несет около двух новых мутаций.
Проделанные выше расчеты основаны на частотах возникновения мутаций, обладающих внешним проявлением. В целом по геному темп мутирования составляет не менее 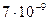 замен на одну нуклеотидную пару в год. У млекопитающих число нуклеотидных пар в диплоидном геноме составляет около
замен на одну нуклеотидную пару в год. У млекопитающих число нуклеотидных пар в диплоидном геноме составляет около 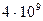 . Следовательно, нуклеотидные замены у млекопитающих происходят с частотой не менее
. Следовательно, нуклеотидные замены у млекопитающих происходят с частотой не менее 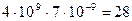 в год на диплоидный геном. Ясно, что мутационный процесс обладает колоссальными возможностями поставлять новый наследственный материал.
в год на диплоидный геном. Ясно, что мутационный процесс обладает колоссальными возможностями поставлять новый наследственный материал.
Важный шаг в генетике популяций был сделан в 1926 году С. С. Четвериковым. Исходя из закона Харди – Вайнберга, С. С. Четвериков доказал неизбежность генетической разнородности природных популяций при том, что новые мутации непрерывно появляются, но остаются обычно скрытыми (рецессивными), а в популяции идет свободное скрещивание.
Из расчетов Четверикова следовало, а впоследствии это было полностью подтверждено практикой, что даже редкие и вредные для особи мутантные гены будут надежно укрыты от очищающего действия естественного отбора в гетерозиготах (организмах со смешанной наследственностью) с доминирующими безвредными генами нормального дикого типа. Это значит, что даже вредная гетерозигота (организм с однородной наследственностью) мутация будет сохраняться в виде генетической «примеси» в течение ряда поколений. Мутация будет как бы поглощена популяцией, из-за чего за внешним однообразием особей одной популяции неизбежно скрывается их огромная генетическая разнородность. Четвериков это выразил так: «Вид, как губка, впитывает в себя гетерозиготные геновариации, сам оставаясь при этом все время внешне (фенотипически) однородным». Для жизни популяций эта особенность может иметь два разных следствия. В огромном большинстве случаев при изменении условий среды вид может реализовать свой «мобилизационный резерв» генетической изменчивости не только за счет новых наследственных изменений у каждой особи, но и благодаря «генетическому капиталу», доставшемуся от предков. Благодаря такому механизму наследования популяция приобретает пластичность, без чего невозможно обеспечить устойчивость приспособлений в меняющихся условиях среды. Однако изредка возможен и другой исход: редкие скрытые вредные мутации иногда могут встретиться у потомства совершенно здоровых родителей, приводя к появлению особей с наследственными заболеваниями. И это – тоже закономерное, неистребимое биологическое явление, своего рода жестокая плата популяции за поддержание своей наследственной неоднородности.
С. С. Четверикову популяционная генетика обязана еще одним открытием, которое было изложено в маленькой, всего на 4 страницы, заметке «Волны жизни», опубликованной в 1905 году на страницах «Дневника Зоологического отделения Императорского общества любителей естествознания и этнографии» в Петербурге. Он обратил внимание, что поскольку любая природная популяция имеет конечную, ограниченную численность особей, это неизбежно приведет к чисто случайным статистическим процессам в распространении мутаций. При этом популяции всех видов постоянно меняют численность (численность грызунов в лесу может от года к году изменяться в сотни, а многих видов насекомых – в десятки тысяч раз), из-за чего в разные годы распространение мутаций в популяциях может идти совершенно по-разному. От громадной популяции птиц, насекомых, зайцев и других животных в трудный для переживания год может остаться всего несколько особей, причем иногда совершенно нетипичных для бывшей популяции. Но именно они дадут потомство и передадут ему свой генофонд, так что новая популяция по составу генетического материала будет совершенно иной, чем прежняя. В этом проявляется генетический «эффект основателя» популяции. Постоянно изменяется и геном в популяциях человека. К. Альстрем на материале в южной Швеции показал, что в популяции человека передается следующему поколению далеко не весь имеющийся генофонд, а лишь избранная, а то и случайно «выхваченная» часть. Так, 20% поколения здесь вовсе не оставили потомков, зато 25% родителей, которые имели трех и более детей, дали 55% численности следующего поколения.
Постоянное давление мутаций и миграции генов, а также выщепление биологически менее приспособленных генотипов по сбалансированным полиморфных локусам создает проблему так называемого генетического груза.
Понятие генетического груза ввел Г. Меллер в 1950 году в работе «Наш груз мутаций». По его расчетам, от 10 до 50% гамет у человека содержат хотя бы одну вновь возникшую мутацию. Слабо вредящие мутации, если только они проявляются в гетерозиготе, способны нанести популяции больший урон, чем полностью рецессивные летальные мутации. Каждый из нас является носителем по крайней мере восьми вредных мутаций, скрытых в гетерозиготном состоянии. Г. Мёллер в соавторстве с Н. Мортоном и Дж. Кроу [1956] произвели оценку генетического груза мутаций путем сравнения детской смертности в случайных выборках из популяций и в семьях, где имели место браки между родственниками. Они выделили собственно мутационный груз, возникающий в результате мутационного давления, и сегрегационный груз как следствие расщепления. Ими предложены расчеты летального эквивалента, соответствующего числу мутаций, дающих вместе летальный исход. Так, один летальный эквивалент может соответствовать одной летальной мутации, двум полулетальным и т. д. Было показано, что средняя величина генетического груза у человека равна 3–5 летальным эквивалентам.
Ю. П. Алтухов с коллективом сотрудников [1989] в результате длительного изучения локальных стад рыб – больших изолированных друг от друга популяций с исторически сложившейся субпопуляционной структурой – пришел к выводу о высокой их устойчивости во времени и пространстве. Изменчивость на уровне отдельных субпопуляций не играет самостоятельной роли и отражает локальные различия действия отбора в силу гетерогенности условий обитания, а также влияние случайных факторов. К аналогичному заключению еще раньше пришел Ю. Г. Рычков при исследовании с сотрудниками изолированных групп популяций человека – коренного населения циркумполярной зоны Евразии. Американский генетик и селекционер И. М. Лернер еще в 1954 году выдвинул представление о генетическом гомеостазе, определив его как способность популяции приводить в равновесие свою генетическую структуру и противостоять внезапным изменениям. Один из важных механизмов генетического гомеостаза – отбор в пользу гетерозигот, ведущий к сбалансированному равновесию. Вместе с тем этот же механизм служит причиной образования генетического груза – выщепляющихся гомозиготных классов особей. Такой груз был назван сбалансированным и рассматривается как плата за поддержание гетерозигот, причисленных к генетической элите популяции.
Частоты генов в популяциях. Для описания ситуаций в популяционной генетике создано несколько математических моделей. Важнейшая заслуга в разработке этой области принадлежит Сьюэллу Райту. Еще в 1928 году Валунд [Wahlund, 1928] установил, что если большая популяция подразделена на K панмиктических групп, то в такой совокупности наблюдается эффект, подобный последствиям инбридинга в неподразделенной популяции: доля гомозигот возрастает на величину межпопуляционной вариансы частот генов за счет уменьшения доли гетерозигот.
Принципиальный вклад в описание локальной дифференциации частот генов в подразделенной популяции в терминах F - статистики был внесен С. Райтом [1943, 1951], который обосновал несколько F - коэффициентов как показателей меры генетической дифференциации:
1) FIT – коэффициент инбридинга особи относительно целой (T) популяции;
2) FIS – коэффициент инбридинга особи относительно субпопуляции (S);
3) FST – коэффициент инбридинга субпопуляции относительно всей подразделенной популяции.
Соотношение между этими величинами задается равенством:
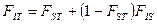 .
.
Коэффициент FST был предложен С. Райтом еще в 1943 году и с тех пор неоднократно использовался при анализе распределений частот генов в природных подразделенных популяциях. Коэффициент Райта представляет большой интерес, так как позволяет вычленить некоторые важные влияния популяционной подразделенности на генетическую структуру. Для этой цели С. Райт предложил две оригинальные модели популяций: «островная модель» и «изоляция расстоянием».
Островная модель. Известны для варианта островной модели популяции: 1) подразделенность вида на множество свободно скрещивающихся внутри себя субпопуляций генетически эффективного объема N, каждая из которых с равной вероятностью и с одинаковой интенсивностью m обменивается генами с любой другой; 2) большая панмиктическая популяция («материк»), окруженная множеством изолированных, генетически дифференцированных малых колоний («острова»), каждая из которых получает гены с «материка» с интенсивностью m на поколение. Эффектами обратной миграции можно пренебречь.
Мерой случайной дифференциации субпопуляций в такой системе служит межгрупповая варианса генных частот:
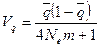 ,
,
и, следовательно, условие равновесия между дрейфом и миграцией генов в терминах FST – статистики может быть записано как:
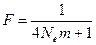 .
.
Более строгое решение относительно Vq задается формулой (Wright, 1943):
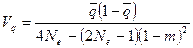 .
.
Как следствие взаимодействия дрейфа и миграции мы имеем вероятностное распределение частот генов. В любой момент времени T оно представляет функцию от 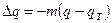 как меры систематического давления миграции и
как меры систематического давления миграции и 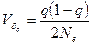 – выборочной вариансы частоты гена в одном поколении за счет изоляции, т. е. случайного дрейфа (Wright, 1938):
– выборочной вариансы частоты гена в одном поколении за счет изоляции, т. е. случайного дрейфа (Wright, 1938):
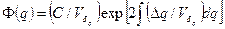 .
.
Если мы обозначим через qi частоту гена в i-й группе 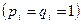 , а через q – частоту этого же гена в подразделенной популяции как в целом, то характерные для нее средняя частота гена и ее варианса будут:
, а через q – частоту этого же гена в подразделенной популяции как в целом, то характерные для нее средняя частота гена и ее варианса будут:
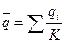 ,
, 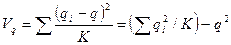 .
.
Соответственно частоты зигот (генотипов) равны
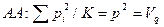
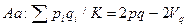
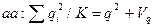
Сопоставляя частоты генотипов с их частотами в популяции, характеризующейся коэффициентом инбридинга F, получим соотношение
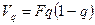 или
или 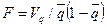 .
.
Поскольку величина F характеризует подразделенную популяцию в целом, то соответствующие частоты генотипов в ней равны частотам, которые были бы свойственны отдельной инбредной популяции. Иными словами, «подразделенность популяции на отдельные скрещивающиеся группы формально эквивалентна наличию инбридинга во всей популяции» [Ли, 1978].
Общая формула стационарного распределения частот генов в островной модели представляет b-функцию плотности вероятности следующего вида:
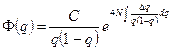 ,
,
где С – нормирующий множитель, подбираемый так, чтобы
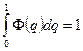 ,
, 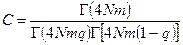 .
.
В зависимости от того, каким сочетанием случайных и систематических факторов задается распределение, оно принимает вид:
а) при уравновешивании случайного дрейфа генов их миграцией
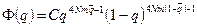 ,
,
где p и q– частоты аллелей в субпопуляциях; 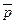 и
и 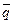 – средние частоты аллелей для подразделенной популяции как целого; N – эффективный размер популяции; m – коэффициент миграции;
– средние частоты аллелей для подразделенной популяции как целого; N – эффективный размер популяции; m – коэффициент миграции;
б) при объединенном эффекте воздействия изоляции, миграции и отбора:
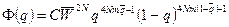 ,
,
где все обозначения те же, что и в предыдущем выражении, а 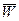 – средняя внутрилокусная приспособленность популяции, определяемая через суммирование приспособленностей генотипов с учетом их частот.
– средняя внутрилокусная приспособленность популяции, определяемая через суммирование приспособленностей генотипов с учетом их частот.
Стационарные распределения могут описывать: 1) распределения частот аллелей многих локусов в одной и той же популяции в случае нейтральности или при примерно одинаковом давлении отбора на каждый локус; 2) распределение генных частот какого-либо локуса в последовательных поколениях одной и той же стационарной популяции; 3) распределение частот аллелей одного или нескольких локусов в совокупности полностью или частично изолированных популяций. Все три типа распределений математически эквиваленты.
В островной модели величина коэффициента миграции генов не зависит от степени удаленности популяций. Райт [Wright, 1943] и Малеко [1955, 1957] математически исследовали ту же популяцию, в которой интенсивность обмена между субпопуляциями зависит от расстояния. Эта модель носит название «изоляция расстоянием» и предполагает популяцию, непрерывно распределенную на большой территории, существенно превышающей радиус индивидуальной активности особи в репродуктивный период. Особенности локальной дифференциации в такой системе зависят от репродуктивной величины, или «соседства», откуда случайно происходят родители, а также от размерности ареала.
Согласно С. Райту [1951], размер соседства приблизительно соответствует числу генетически эффективных особей внутри круга, радиус которого равен удвоенному стандартному отклонению s протяженности миграции в одном направлении в данном поколении (или – что то же – дистанции между местами рождения родителей и потомков).
Дифференциация очень велика, когда 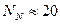 , намного меньше, но все же достаточно выражена при
, намного меньше, но все же достаточно выражена при 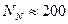 и почти соответствует панмиксии, когда
и почти соответствует панмиксии, когда 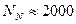 .
.
М. Кимура [1953] предложил другую популяционную модель популяционной структуры. Она носит название «лестничная модель» и представляет ситуацию, промежуточную между райтовской островной моделью и моделями непрерывно распределенных популяций Райта и Малеко.
Лестничная структура миграции генов. В этой модели, как и в островной, рассматривается совокупность колоний, однако обмен особями происходит только между соседними колониями, и, таким образом, непосредственно зависит от удаленности колоний друг от друга.
При равновесии межпопуляционная варианса генных частот
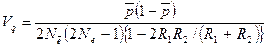 ,
,
где 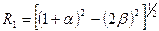 ;
; 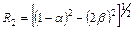 , в которых
, в которых 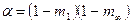 и
и 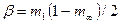 ; в этих уравнениях m1 – интенсивность миграций между смежными колониями, а
; в этих уравнениях m1 – интенсивность миграций между смежными колониями, а 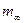 – давление миграции генов извне на всю совокупность колоний (соответствует коэффициенту m в островной модели С. Райта). Когда m1= 0, то
– давление миграции генов извне на всю совокупность колоний (соответствует коэффициенту m в островной модели С. Райта). Когда m1= 0, то 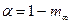 ,
, 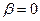 , и выражение сводится к формуле С. Райта. Островная модель С. Райта, таким образом, представляет собой частный случай лестничной модели в отсутствие обмена генами между соседними колониями.
, и выражение сводится к формуле С. Райта. Островная модель С. Райта, таким образом, представляет собой частный случай лестничной модели в отсутствие обмена генами между соседними колониями.
Важнейшая особенность подразделенности, также исследованная теоретически, – способность подразделенных популяций поддерживать значительно большее генетическое разнообразие в сравнении с панмиктическими популяциями сопоставимого размера. Считается, что именно такое разнообразие и позволяет подразделенной популяции более эффективно реагировать на изменения среды и вслед за ними изменять свою генотипическую структуру – тезис, играющий решающую роль в эволюционной концепции С. Райта, известной под названием «теория смещающегося равновесия», в которой «поверхность» изображается топографической картой с вершинами и долинами на едином ландшафте генных комбинаций. В этой модели важнейшее заключение состоит в том, что «эволюционный процесс зависит от постоянно смещающегося баланса между факторами стабильности и изменений и что наиболее благоприятное условие для этого – наличие тонко подразделенной структуры, в которой изоляция и перекрестная коммуникация поддерживаются в соответствующем равновесии» [Wright, 1951].
Дата добавления: 2015-04-11; просмотров: 4353;