Современное понимание механизмов общей анестезии
Выяснение сложных механизмов действия общих анестетиков на процессы, связанные с формированием и распространением возбуждения в нервной системе, несмотря на резко возросшие в последние десятилетия методические возможности, связано с большими трудностями. Сложившееся представление об этих механизмах предусматривает влияние анестетиков прежде всего на процессы образования и распространения потенциала действия на уровне самих нейронов и, особенно, межнейронных контактов. Поскольку синапсы являются в рефлекторной цепи звеном не только наиболее сложным, но и наиболее подверженным влиянию на его функцию различного рода эндогенных и экзогенных факторов, тормозящее влияние анестетиков на передачу импульсов в них проявляется больше, чем на возбудимость мембраны нейрона.
Предположение о том, что общие анестетики реализуют свое специфическое действие в основном через синапсы, впервые высказал в 1906 г. известный английский физиолог Ch. Sherrington. В дальнейшем было установлено, что общие анестетики оказывают выраженное тормозящее действие на синаптическую передачу в дозах, которые существенно не влияют на распространение возбуждения по мембране нейрона. Для торможения распространения потенциала действия на мембране необходимы более значительные концентрации анестетика, хотя сам механизм торможения в том и другом случае аналогичен.
Механизм угнетения возбудимости нейронов и торможения синаптической передачи возбуждения под влиянием анестетиков полностью не раскрыт. При объяснении его некоторые исследователи исходят из того, что молекулы анестетика образуют на мембране нейрона своеобразный плащ, затрудняющий прохождение через нее ионов и, следовательно, препятствующий процессу деполяризации мембраны. Другое представление основано на изменении под влиянием анестетиков функции катионных «каналов» клеточных мембран [Mullins L.J., 1975; Kendic J.J., Tradell J., 1976]. Эти авторы допускают два варианта такого рода блокады. Первый из них сводится к перекрытию молекулами анестетика входа в «каналы», предназначенные для транспорта Na+. Второй вариант предусматривает такие изменения в структуре липидов мембраны или белковых ее элементов, которые лишают натриевые «каналы» способности раскрываться в ответ на возбуждение (рис. 6.1). Есть предположение, что в этом процессе играют значительную роль Са2+, которые под влиянием анестетиков теряют подвижность и задерживаются в мембране.
Тот факт, что тормозящее влияние общих анестетиков на распространение возбуждения больше проявляется в синапсах, чем в пределах самого нейрона, вызвал интерес ряда исследователей. В этой области накоплены некоторые данные [Шаповалов А.И., 1966; Gage P.W., Hammil О.R., 1981], свидетельствующие о вероятности действия анестетиков на различные звенья синаптической передачи возбуждения (рис. 6.2). Выяснено, что торможение передачи возбуждения под влиянием некоторых анестетиков начинает проявляться на подходе к пресинаптической мембране. Здесь, в терминальной части нервного волокна, анестетики снижают гарантийный фактор потенциала действия. В самом синапсе влияние анестетиков может проявляться угнетением образования медиатора, снижением чувствительности к нему рецепторов пресинаптической и постсинаптической мембран [Alper M., Flock W., 1969; Franks N., Lieb W., 1982; Souter S. Q. et al., 1980]. В результате уменьшается возможность формирования постсинаптического возбуждающего потенциала, необходимого для распространения возбуждения. Однако и в тех случаях, когда постсинаптический потенциал приводит к формированию потенциала действия, при небольшом гарантийном факторе он может гаснуть в результате действия анестетика на терминаль нервного волокна нейрона, воспринимающего возбуждение.
Естественно, что различные анестетики неодинаково влияют на основные функциональные звенья синапсов. Это связано как с неоднородностью последних, так и с особенностями свойств отдельных анестетиков. Одни из них тормозят передачу возбуждения преимущественно на уровне терминалей нервных волокон, другие — путем снижения чувствительности рецепторов мембран к медиатору или угнетения его образования.
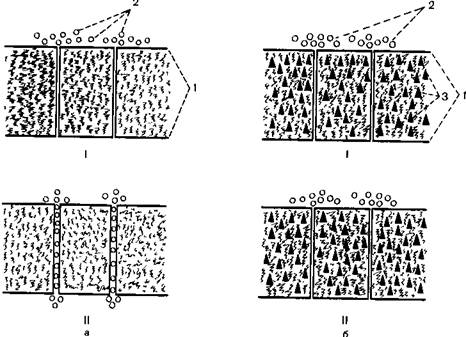
Рис. 6.1.Натриевые каналы мембраны нейрона в физиологических условиях (а), при насыщении мембраны общим анестетиком (б).
I - до возбуждения, II — под влиянием потенциала действия, 1 — мембрана нейрона 2 — ионы Na+ ; 3 — молекулы анестетика.
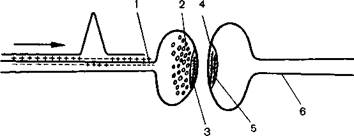
Рис. 6.2.Вероятные зоны действия общих анестетиков на симпатическую передачу возбуждения J
1 - пресинаптическая терминаль нервного волокна, 2 — зона образования медиатора 3 рецепторы пре синаптической мембраны, 4 рецепторы почсинаптической мембраны, 5 зона формирования постсинап тического потенциала действия 6 терминаль постсинаптического нервного волокна.
Косвенным подтверждением преимущественного действия общих анестетиков в зоне межнейронных контактов может служить функционирование недавно открытой антиноцицептивной системы организма. Она, в современном понимании, представляет собой совокупность механизмов, регулирующих болевую чувствительность и оказывающих тормозящее влияние на ноцицетивную импульсацию в целом. Больше других к настоящему времени изучен антино-цицептивный механизм, функционирующий на основе взаимодействия имеющихся в центральной нервной системе специфических рецепторов с оделенными опиатами [Игнатов Ю.Д., 1984; Калюжный Л.В., 1984; Лиманский Ю.П., 1986]. Поскольку по химической структуре последние близки морфину, эти рецепторы, взаимодействуя с экзогенными опиатами, также дают анальгетический эффект. Предполагают, что он возникает в результате торможения ноцицеп-тивной импульсации на уровне синапсов спинного и головного мозга, т.е. на основе такого же механизма, который считают характерным и для наркотических веществ.
Несмотря на то что рассмотренное выше понимание механизма действия общих анестетиков еще не полностью обосновано фактическими данными по сравнению с разработанными ранее теориями наркоза, эта концепция представляется более целостной и в большей мере соответствует современному состоянию нейрофизиологии.
Однако при всей ценности сведений об интимном механизме действия анестетиков на электрические процессы в мембране, с которыми связаны формирование и распространение возбуждения, они не раскрывают сущности общей анестезии как своеобразного функционального состояния нервной системы организма. Поскольку в клинических условиях используемая концентрация анестетиков не вызывает полной ареактивности нейронов и блокады синаптической передачи, а лишь оказывает тормозящее влияние на их функцию, важно знать направление и последовательность функциональных изменений в зависимости от насыщения нервной системы наркотическими веществами. Дать научно обоснованный ответ на возникшие вопросы оказалось очень сложно. Существенный вклад в выяснение характера нейрофизиологических процессов, связанных с действием на организм наркотических веществ, внесли Н.Е. Введенский (1901), А.А. Ухтомский (1933), В.С. Галкин (1955) и др.
Особого внимания заслуживает объяснение влияния общих анестетиков на функцию нервной системы, которое дал и в значительной степени научно обосновал Н.Е. Введенский. Исходя из разработанной им теории парабиоза, он пришел к заключению, что наркотические вещества действуют на нервную систему как сильные раздражители и, подобно последним, вызывают соответствующие фазы парабиоза, которые характеризуются последовательным снижением физиологической лабильности отдельных нейронов и нервной системы в целом. При определенном для каждого анестетика уровне насыщения мозга снижение лабильности механизмов, лежащих в основе формирования и распространения нервных импульсов, достигает такой степени, которая обусловливает торможение функций ЦНС, проявляющееся клинически состоянием наркоза. В дальнейшем эта концепция получила развитие в трудах А.А. Ухтомского и некоторых других отечественных ученых.
В аспекте рассматриваемой концепции заслуживают внимания научные факты, полученные Т. Biscoe и R. Millar (1966) при выяснении влияния наркотических веществ на проведение вызванных импульсов различной частоты. Авторы установили, что на фоне одинакового насыщения общими анестетиками блокирующий эффект их тем значительнее, чем больше частота наносимых раздражений.
Концепция об изменении под влиянием наркотических веществ физиологической лабильности нейронов и, особенно, синапсов позволила приблизиться к пониманию того, что в каждый данный момент общей анестезии степень торможения функции различных отделов мозга оказывается неодинаковой. Этот феномен может быть объяснен неодинаковой исходной функциональной лабильностью отделов мозга. Такое понимание нашло убедительное подтверждение в том, что наряду с корой больших полушарий наиболее подверженной тормозящему влиянию наркотических веществ оказалась функция ретикулярной формации, это явилось предпосылкой для разработки ретикулярной теории наркоза.
Развитию ретикулярной теории способствовали два важных научных факта, установленных при изучении функции ретикулярной формации: 1) она играет активирующую роль в отношении лежащих выше отделов ЦНС; 2) разрушение определенных зон ретикулярной формации вызывает состояние, близкое к медикаментозному сну или наркозу. Это привело к заключению о вероятной связи специфического действия наркотических веществ с влиянием их прежде всего на функцию ретикулярной формации. Экспериментальные исследования показали, что на фоне действия некоторых общих анестетиков не возникает характерная для обычных условий десинхронизация биотоков мозга в ответ на электрическое раздражение ретикулярной формации. Этот факт, установленный впервые G. Moruzzi и Н. Magoun (1949), в дальнейшем был подтвержден другими исследователями.
Таким образом, сформировалось представление о том, что эффект общих анестетиков является результатом торможения рефлекторных процессов на уровне сетевидной субстанции мозга. При этом устраняется восходящее активизирующее влияние ее, что приводит к деафферентации вышележащих отделов ЦНС.
Несмотря на убедительные данные в пользу ретикулярной теории наркоза, которая в настоящее время получила широкое признание, результаты некоторых исследований свидетельствуют о том, что она не может быть признана универсальной. В частности, Т.М. Дарбинян и В.Б. Головчинский (1976) на основании экспериментальных данных показали, что при наркотизации диэтиловым эфиром и циклопропаном торможение функции мозга распространяется не от ретикулярной формации вверх, а в соответствии с ранее сложившимся представлением, т.е. сверху вниз.
Помимо этого, заслуживают внимания сообщения о том, что при общей анестезии барбитуратами более раннее торможение ретикулярной формации по сравнению с корой может быть результатом влияния анестетика не столько на нее, сколько на структуры спинного мозга, обеспечивающие афферентный поток импульсов [Белоярцев Ф.Ф., 1974].
Таким образом, со времени открытия наркоза и признания большого значения его в обеспечении условий для оперативных вмешательств учеными, теоретиками и клиницистами затрачено много усилий с целью выяснения механизма действия наркотических веществ и раскрытия физиологической сущности общей анестезии в целом. В этой области достигнуто очень многое. Однако проблема оказалась чрезвычайно сложной. Многие относящиеся к ней вопросы остаются без убедительного ответа.
СПИСОК ЛИТЕРАТУРЫ
Анохин П.К. Современное представление о физиологических механизмах центрального действия наркотиков // Общее обезболивание в хирургии.— М., 1955.— С. 28—66.
Белоярцев Ф.Ф. Компоненты общей анестезии.— М.: Медицина, 1974.— С. 114—130.
Введенский Н.Е. Возбуждение, торможение и наркоз 1901 г. // Избранные произведения.— М., 1951.— Т. 4.
Дарбинян Т.М., Головчинский В.Б. Механизмы наркоза.— М.: Медицина, 1972.
Галкин В.С. Краткий очерк теории наркоза: Лекции для врачей.— Л., 1953.
Калюжный Л.В. Физиологические механизмы регуляции болевой чувствительности. — М.: Медицина, 1984.
Лиманский Ю. П. Физиология боли.— Киев: Здоров'я, 1986.
Ухтомский А.А. Физиологический покой и лабильность как биологические факторы - Собр соч.—Л , 1952.— Т. 4.
Albrecht R. F., Miletich D. J Speculations on the molecular nature of anaesthesia // Gen. Pharmacol.- 1988. - Vol. 19, N 3. - P. 339 -346.
Alper M., Flack W The peripheral effects of anesthetics//Amer. Rev. Pharm. 19(59 Vol 9 P. 274 279
Biscoe Т., Millar R. The effect of cyclopropane, halothane and ether on simpathetic ganglionic transmission // Brit. J. Anaesth.— 1966.— Vol. 38.- P. 3—9.
Franks N.. Lieb W. Molecular mechanism of general anesthesia // Nature.— 1982. - Vol 300.— P. 487—493.
Gage P. W., Hammil O.P. Effects of anesthetics on ion channels and synapses // Intern. Rev. Physiol/Ed. R. Porter.—Baltimore, 1981.—Vol. 25.—P. 1—45. Hodgkin A., Katz B. The effect of sodium ions on the electrical activity of the giant axon of the squid//J. Physiol.— 1949.-Vol. 108.—P. 37—45.
Halsey M. J. Anaesthetic mechanisms//Brit. J. Hosp. Med.-— 1986.—Vol. 36.—N 6.—P. 445— 447.
Kendig J.J., Tradell J. Approaches to a theory of anaesthetic action // Scientific foundation anaesthesia/Ed. C. Feldman.— London, 1976.— P. 280—288.
Kendig J.J., Grossman Y., Maclver M. B. Pressure reversal of anaesthesia: a synaptic mechanism // Brit. J. Anaesth.— 1988.—Vol. 60, N 7.— P. 806—816.
Maclver M.В., Roth S.H. Anaesthetics produce differential actions on the discharge activity of a single neuron // Europ. J. Pharm.— 1987.— Vol. 139, N 2.— P. 43—52.
Maclver M. В., Roth S. H. Inhalation anaesthetics exhibit pathway-specific and differential actions on hippocampal synaptic responses in vitro // Brit. J. Anaesth.— 1988.— Vol. 60, N 6.— P. 680—691.
Moruzzi G., Magoun H. Brain stem reticular formation and activation of the EEG // Electroencephal. clin. Neurophysiol.— 1949.—Vol. 1.— P. 455—473.
Mullins L. J. Anesthesia // Molecular mechanisms of anesthesia/Ed. B. Fink.— New York, 1975.— P. 237—243.
Roth S. H. Physical mechanisms of anesthesia//Ann. Rev. Pharm. Toxicol.— 1979.— Vol. 19.— P. 159—178.
Sauter S. G., Elliot /., Raftery M. A. Actions of anethetics and high pressure on cholinergic membranes. Molecular mechanisms of anesthesia // Progress in anesthesiology/Ed. B. P. Fink. New York, 1980.—Vol. 2.—P. 199—207.
Trudell J. R. Molecular Basis for unitary theories of inhalation anaesthesia // Inhalation anaesthesia today and tomorrow/Ed. K. Peter, F. Jesch.— New York, 1982.— P. 45—53.
Дата добавления: 2015-03-09; просмотров: 942;