Запечатление, или импринтинг
Импринтинг исследовали главным образом у выводковых и зрело-рождающихся видов, поскольку многие элементы поведения, "уже готовые" сразу после рождения, позволяли более надежно оценивать реакции животных. Однако сходные с импринтингом явления наблюдались и у незрелорождающихся животных, например реакция следования за матерью, хорошо известная у копытных, землероек и даже у бурого медведя (см.: Хайнд, 1975).
Запечатление считается специфическим видом обучения, очень важным для формирования в онтогенезе индивидуальных адаптации (см: гл. 3). Оно заключается в том, что молодое животное или птица начинает следовать за тем животным (или предметом), которые попали в поле его внимания в первые часы после появления на свет. Как говорит об этом Д. Мак-Фарленд (1988), импринтинг – это "...совершенно неразборчивая привязанность к движущимся объектам".
Как известно, импринтинг начали изучать, наблюдая реакцию следования птенцов выводковых птиц (из отрядов куриных и гусеобразных) за матерью. Одним из первым описание импринтинга дал английский ученый Д. Сполдинг (см.: 2.3).
К. Лоренцу принадлежит заслуга подробного и тщательного описания импринтинга на основе наблюдений за дикими гусями, выращенными человеком. Он выделил две важные особенности, которые отличают данный феномен от других типов обучения. Во-первых, этот феномен приурочен к определенному периоду онтогенеза, называемому чувствительным (или критическим). У разных видов животных и птиц он имеет разную длительность. Во-вторых, процесс импринтинга необратим. Если запечатление объекта молодой птицей или зверем произошло, то эффект этого раннего опыта практически устранить нельзя. Например, если у птенца произошло запечатление на особь другого вида, позднейшие его контакты с представителями своего вида не смогут изменить возникшей привязанности.
Считается, что существует импринтинг двух типов – запечатление привязанности и половое запечатление. Хотя такое подразделение несколько условно, два типа импринтинга, тем не менее, следует анализировать с помощью разных подходов.
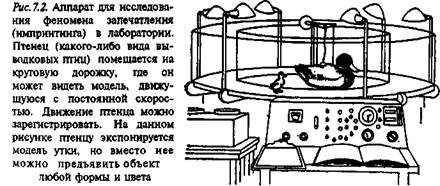
Запечатление привязанностиобычно исследуют, анализируя этоло-гические и физиологические закономерности собственно реакции следования. Наиболее часто для этого используют птенцов выводковых птиц (рис. 7.2 и 7.3).
Специальные эксперименты были проведены американским исследователем Э. Хессом (Hess, 1964). Вкратце суть приемов анализа запечатления такова. Появление птенцов из яйца происходит в темноте. Впервые они получают зрительный опыт на экспериментальной площадке, куда их можно помещать в том или ином возрасте, и где на специальной круговой дорожке им демонстрируют тот или иной объект. Объект может перемещаться с разной скоростью, так что реакция следования за ним у экспериментального птенца может быть охарактеризована количественно. Движущимся предметом может быть чучело птицы, коробка или что-либо еще. Чем ярче этот предмет окрашен, тем более сильную реакцию следования он вызывает. Упрочению реакции способствуют также звуки, которые сопровождают перемещение предмета.
|
Более подробное исследование показало, что сенсорные раздражители, которые воспринимает молодая птица, вызывают запечатление, имеющее разную "стойкость". Так, запечатление "природных" раздражителей, или "стимул-объектов", т.е. тех объектов, с которыми данный вид сталкивается в естественной среде обитания, происходит быстрее, чем искусственных. Это заставляет предполагать существование генетически детерминированной "предуготованности" к запечатлению определенных стимулов. Кроме того, запечатление искусственных, лабораторных объектов иногда оказывается непрочным. Опыты Хесса показали, что утята, у которых произошло запечатление на человека (контакт длился 20 часов непрерывно), все-таки предпочитали двигаться за живой уткой даже после короткого периода (1,5 часа) контакта с ней.
Исследование гнездового поведения уток показало, что еще до вылупления между матерью и птенцом происходит активный обмен акустическими сигналами. Это, видимо, является одним из факторов, благодаря которым импринтинг естественных стимул-объектов оказывается более прочным: маленькие утята оказываются уже "настроенными" на определенные аудиометрические характеристики материнского сигнала.
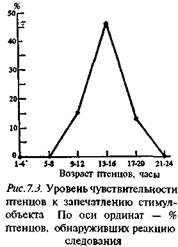
Данные лабораторных экспериментов подтвердили первоначальное положение Лоренца (см. гл. 3) о том, что существует определенный период после вылупления птенца (или рождения детеныша), в течение которого запечатление может произойти. У утят готовность к запечатлению достигает максимума в период от 13 до 16 часов жизни (см. рис. 7.3). В более ранние и в более поздние сроки предъявление такому птенцу стимул-объектов оказывается менее эффективным. Такие же периоды повышенной чувствительности, играющие важную роль в формировании поведения взрослой особи (см. ниже), обнаруживаются и у млекопитающих.
В успешности запечатления существенную роль играет специфическая зрительная активация. В лабораторных экспериментах было показано, что у цыплят, подвергнутых действию света сразу после вылупления, реактивность на предъявленные им предметы выше, чем у цыплят, содержавшихся в темноте. При содержании цыпленка в изоляции реактивность его в отношении объектов запечатления сохраняется дольше по сравнению с цыплятами, которые содержались в группах. Этот феномен объясняют тем, что в группе цыплят происходит запечатление друг друга. Наблюдения в природе подтверждают, что, например, у древесной кряквы птенцы покидают гнездо в момент пика чувствительного периода, а взаимное "имп-ринтирование" обеспечивает их безопасность. Если непосредственно за матерью следует только часть выводка, то остальные птенцы бегут друг за другом.
В опытах английского ученого П. Бэйтсона (Bateson, 1966) было показано, что импринтинг у цыплят проходил более успешно, если объект запечатления имел ту же окраску, что и стены помещения, где они содержались до начала опыта. Полагают, что в физиологическом механизме импринтинга определенную роль играет способность птенца отличить знакомое от незнакомого. Значительная роль в работе такого механизма принадлежит видоспецифической и генетической предрасположенности к запечатлению, а также, возможно, пренатальному опыту (выше уже упоминалось о роли звуковых сигналов утки-матери в запечатлении ее облика).
Считается, что сам факт осуществления запечатления ведет к окончанию чувствительного периода, которое совпадает с началом формирования у птенца реакции страха. Это совпадение не случайно. У молодых животных и птиц состояние страха возникает при встрече их с незнакомыми объектами, и в противоположность тому, как это бывает при запечатлении, они стремятся избегать новых предметов. Можно полагать, что в основе такой "реципрокности" лежит какое-то общее звено нервных механизмов обоих процессов – запечатления и реакции избегания.
Половой импринтинг– это явление, выражающееся в том, что данное взрослое животное адресует свое половое поведение особям, контакт с которыми произошел у него в раннем периоде онтогенеза. Эксперименты показали, что запечатление привязанности и половое запечатление не идентичны и различаются, по меньшей мере, сроками наступления чувствительных периодов. Половой импринтинг исследуют как у выводковых, так и у птенцовых видов птиц. Тестирование предпочтения полового партнера проводят по достижении подопытной птицей зрелого возраста.
Для исследования полового импринтинга птицу выращивают либо с особями своего вида, но отличающимися по окраске (такие эксперименты проводили со снежными гусями, вьюрками, голубями), либо с особями других видов. У самцов и самок индюков отмечали формирование полового импринтинга на человека. Немецкий исследователь К. Иммельман в опытах с вьюрками помещал одно яйцо, например зебровой амадины, в кладку другого вида – бенгальской амадины. Воспитанный таким образом самец зебровой амадины по окончании гнездового периода содержался в изоляции вплоть до наступления половой зрелости. При тестировании такой самец все ухаживания адресовал самке бенгальской амадины, т.е. виду, к которому принадлежали вырастившие его птицы.
Половой импринтинг обнаруживает некоторую степень "обратимости". Если такого усыновленного самца принудительно соединить с самками своего вида, то в конце концов спаривание произойдет, и самец будет принимать участие в воспитании потомства. Однако если самцу еще раз дать возможность выбрать самку, то он опять выберет самку, принадлежащую к виду его приемных родителей. Хрестоматийный пример такого аномального полового запе-чатления – это случай, когда самец зебровой амадины адресовал ухаживание руке человека, которая его "вырастила", и игнорировал самок своего вида (рис. 7.4). У гусей и уток самцы и самки видов-воспитателей "запечатлеваются" воспитанниками по разному, причем рационального объяснения этому феномену пока нет.
Феномены, во многом сходные с запечатлением, обнаружены также у млекопитающих. Наиболее отчетливо их можно видеть у зрелорождающихся видов (по аналогии с выводковыми птицами). Это, например, копытные, а среди грызунов – морские свинки. Только что родившийся ягненок (или козленок) будет следовать за человеком, если тот начал поить его из соски, а детеныш не имел контакта с матерью. Даже во взрослом состоянии такое животное, попав в стадо, будет реагировать на этого человека особым образом – упорно следовать за ним.

Таким образом, поведение взрослой особи, точнее ее взаимоотношение с сородичами, оказывается сильно видоизмененным из-за особенностей ее контактов в короткий период раннего онтогенеза.
Реакция следования как одна из важнейших адаптации раннего онтогенеза описана у бурого медведя в уникальных опытах B.C. Па-жетнова по выращиванию медвежат-сирот (Пажетнов, 1991).
В формировании поведения млекопитающих важная роль принадлежит обонянию, и в развитии запечатления привязанности важны обонятельные сигналы, полученные в раннем возрасте. Они определяют поведение взрослого животного. Показано, например, что для мышей, крыс и морских свинок привлекательнее такие половые партнеры, чей запах больше похож на запахи их гнезда.
Животноводы знают, что овца или коза, не имевшие возможности хотя бы короткого контакта с детенышем сразу после рождения, не проявят по отношению к нему материнской заботы. В некоторых случаях это даже сопровождается торможением лактации.
Запечатление привязанности и сходные с ним явления участвуют в формировании общественного поведения млекопитающих. Лучше всего изучены особые чувствительные периоды, важные для последующей "социализации", у хищных, в особенности у домашней собаки. Существование таких периодов давно известно всем собаководам-практикам и любителям. Первой обобщающей работой по генетике и развитию поведения собак была книга американских исследователей Р. Скотта и Дж. Фуллера (Scott, Fuller, 1965). Нормальные социальные контакты молодой собаки формируются в возрасте от 3 до 10 недель. Если выращивать щенков в изоляции до 14 недель, то в дальнейшем они не в состоянии нормально реагировать на сородичей. Если вспомнить проведенные в павловской лаборатории опыты С.Н. Выржиковского и Ф.П. Майорова (1933) по изолированному воспитанию щенков (см.: 2.5), то очевидно, что в таких случаях нарушенным оказывается поведение в целом, а не только способность таких животных к социальным контактам. Эти животные отличались крайней трусливостью, и выработать у них условные рефлексы, легко формировавшиеся у нормальных собак, было практически невозможно.
Такой же чувствительный период существует и для формирования нормальных контактов собаки с человеком, основа которых также лежит в особенностях видоспецифического общественного поведения этих животных.
Биологические механизмы повышенной чувствительности животных к определенному опыту в раннем онтогенезе (чувствительные периоды для импринтинга) исследуются с помощью различных современных методов.
Феномен чувствительного периода есть частный случай более общего явления – существования стадий развития организма, чувствительных к тому или иному внешнему по отношению к данной развивающейся системе воздействию. Существенно отметить, что имеется большое число экспериментальных данных, из которых следует, что действие ряда факторов на развивающийся организм вызывает у взрослых особей значительные и стойкие изменения поведения. К ним относятся стрессирующие раздражители, обеднение или обогащение условий содержания животных, кратковременное воздействие фармакологическими веществами. Изменения затрагивают способность к обучению, уровень двигательной активности и ориентировочно-исследовательских реакций.
Ранние воздействия влияют и на развитие гормональной сферы млекопитающих, в частности изменение уровней половых гормонов в пренатальный период и сразу после рождения видоизменяет половое поведение у взрослых. Максимальная чувствительность детенышей крыс и мышей к введению половых гормонов бывает в возрасте 1–2 дней. Так, например, если в этом возрасте самку подвергнуть воздействию тестостерона, то происходит так называемая маскулинизация поведения, причем видоизмененным по мужскому типу оказывается не только половое поведение, но и уровень агрессивности таких животных.
Формирование способности к видоспецифической песне (популярный объект изучения относительной роли генов и среды в поведении, см.: 7.3.3) у многих видов птиц также связано с восприятием определенных акустических сигналов в особые периоды специфически повышенной чувствительности к ним.
Многие исследователи считают, что для проявления импринтинга как особого вида обучения очень важны врожденные предрасположенности конкретного вида. Очевидно, что, воспринимая данный стимул-объект, едва вылупившийся птенец его запоминает, т.е. происходит фиксация следа памяти. Индивидуальный опыт такого птенца (история его взаимодействия с внешней средой) поддается экспериментальному контролю: например, птенца можно содержать в темноте и показывать ему только определенные предметы в течение фиксированного времени. С помощью методов, применяемых при исследовании обучения и памяти, удалось идентифицировать участок мозга цыпленка, который активируется в результате одномоментного "акта импринтинга", а также определить условия оптимальной фиксации следа памяти.
В экспериментах, подробно описанных в монографии английского ученого Г. Хорна (Хорн, 1988), было показано, что в процессе импринтинга активируется определенная область переднего мозга цыпленка. Как известно, строение переднего мозга птиц иное, чем у млекопитающих. Из корковых структур у них имеется только гомолог старой и древней коры млекопитающих, а новая кора полностью отсутствует. Полушария переднего мозга птиц заняты образованиями ядерного типа, часть из них гомологична базальным ганглиям млекопитающих, в частности полосатому телу (corpus striatum), и выполняет те же функции, а остальные рассматриваются как функциональные аналоги новой коры.
Стриатум птиц и по морфологическим признакам, и по функции четко подразделяется на несколько отделов. Средняя (т.е. медиальная) и расположенная ближе к основанию мозга часть отдела, называемого медио-вентральный гиперстриатум (МВГ), связана с за-печатлением объекта. Это стало известно из экспериментов, проведенных с использованием ряда методов. Во-первых, проводились опыты по "расщеплению" мозга: та половина мозга, которая выключалась из работы и не получала информации о стимул-объекте, не показывала признаков активации, тогда как в работающем полушарии избирательно возбуждался МВГ. Во-вторых, избирательное участие именно МВГ в фиксации следа памяти о единичном событии – восприятии импринтируемого объекта – было доказано методом радиоавтографии. Животному вводят вещество, меченное радиоактивным изотопом, например 14С-урацилом, а на радиоавтографах (отпечатках срезов мозга) оценивают включение этого вещества в РНК.
Более интенсивная активность клеток, в том числе и нейронов, сопровождается более интенсивным их мечением. Метод 2-дезок-сиглюкозы также основан на накоплении радиоактивного изотопа 14Св ткани мозга: в организм вводится 2-дезоксиглюкоза, которая сначала используется клеткой как глюкоза, но не претерпевает полного метаболизма и накапливается в ней. Радиоактивный изотоп позволяет выявлять такое накопление, которое возрастает с увеличением интенсивности энергетического обмена данной клетки или данной области мозга.
Оба метода достаточно убедительно показали, что именно МВГ – это участок мозга, критически важный для осуществления импринтинга у молодых цыплят. Традиционный метод разрушения этого отдела также показал, что для запечатления необходима целостность МВГ. Спонтанная электрическая активность нейронов МВГ отрицательно коррелировала с активностью приближения к стимул-объекту при запечатлении: чем ближе к объекту запечатления – коробке – находился цыпленок, тем ниже была частота разрядов нейронов. Для нейронов других отделов мозга подобной зависимости не обнаружено. Хорн показал, что у цыплят, которые обучились приближаться к экспериментальному объекту, специфически изменялась структура синапсов МВГ, причем характер изменений был таким же, как и при других формах обучения.
Таким образом, с одной стороны, анализ процесса импринтинга помогает исследовать физиологические и биохимические закономерности обучения и памяти. Сдругой стороны, импринтинг – это феномен, выполняющий важную функцию в жизни особи и вида в целом. Как правило, он наблюдается у видов, для которых привязанность к родителям, к семейной группе,, а также к особи противоположного пола составляет существенный аспект социальной организации. У некоторых видов чувствительный период для полового импринтинга (т.е. запечатления особей своего вида) начинается не сразу после рождения, как это бывает в случае запечатления привязанности, а позже. В работах П. Бэйтсона, проведенных на выводковых птицах разных видов, показано, что особь учится узнавать своих собратьев тогда, когда они уже приобрели признаки молодых особей данного вида (Bateson, 1966; Хайнд, 1975). Впоследствии это облегчает взаимное узнавание особей.
В соответствии с эволюционной теорией, запечатление способствует тому, чтобы родители выращивали только собственное потомство, а спаривание происходило бы лишь между особями одного вида.
7.2.
Дата добавления: 2015-01-10; просмотров: 2747;