Потребность в кислороде у различных видов пресноводных рыб
| Потребность в кислороде (cм2/л) | Виды рыб |
| 7—11 5—6 4 0,5 | Виды рыб холодных и быстрых вод: форель (Salrriio frutta), гольян (Phoxinus phoxinus), подкаменщик (Cottus gobio) Хариус (Thymallus thymallus), пескарь обыкновенный (Gobio gobio), голавль (Zeuciscus ; cephalus), налим (Lota lota) Плотва (Rutilus rutilus), ерш (Acerina cerina) Линь (Tinea tinea) |
Среди водных обитателей значительно количество видов, способных переносить широкие колебания содержания кислорода в воде, близкие к его отсутствию. Это так называемые эвриоксибионты. К ним относятся пресноводные олигохеты (Tubifex tubifex), брюхоногие моллюски (Viviparus viviparus). Очень слабое насыщение воды кислородом из рыб могут выдерживать сазан, линь, караси. Однако многие виды являются стеноксибионтными, т. е. могут существовать только при достаточно высоком насыщении воды кислородом, например радужная форель, кумжа, гольян и др. Многие виды живых организмов способны при недостатке кислорода впадать в неактивное состояние, так называемый аноксибиоз, и таким образом переживать неблагоприятный период.
Дыхание гидробионтов осуществляется как через поверхность тела, так и через специализированные органы — жабры, легкие, трахеи. Нередко покровы тела могут служить дополнительным органом дыхания. У отдельных видов встречается комбинирование водного и воздушного дыхания, например, двоякодышащие рыбы, сифонофоры, дискофанты, многие легочные моллюски, ракообразные Yammarus lacustris и др. Вторичноводные животные сохраняют обычно атмосферный тип дыхания как энергетически более выгодный, и поэтому нуждаются в контактах с воздушной средой. К ним относятся ластоногие, китообразные, водяные жуки, личинки комаров и т. д.
Углекислый газ. В водной среде живые организмы кроме недостатка света, кислорода могут испытывать недостаток доступной СО2, например растения для фотосинтеза. Углекислота поступает в воду в результате растворения СО2 содержащегося в воздухе, дыхания водных организмов, разложения органических остатков и высвобождения из карбонатов. Содержание углекислого газа в воде колеблется в пределах 0,2—0,5 мл/л, или в 700 раз больше, чем в атмосфере. СО2 растворяется в воде в 35 раз лучше кислорода. Морская вода является главным резервуаром углекислого газа, так как содержит от 40 до 50 см3 газа на литр в свободной или связанной форме, что в 150 раз превышает его концентрацию в атмосфере.
Углекислый газ, содержащийся в воде, принимает участие в формировании известковых скелетных образований беспозвоночных животных и обеспечивает фотосинтез водных растений. При интенсивном фотосинтезе растений идет усиленное потребление углекислого газа (0,2—0,3 мл/л в час), что приводит к ее дефициту. На увеличение содержания СО2 в воде гидрофиты реагируют, повышая фотосинтез.
Дополнительным источником СО, для фотосинтеза водных растений является также углекислота, которая выделяется при разложении двууглекислых солей и их переходе в углекислые:
Са(НСО3)2 ® СаСО3 + СО, + Н2О
Малорастворимые карбонаты, которые при этом образуются, оседают на поверхность листьев в виде известкового налета или корочки, хорошо заметной при обсыхании многих водных растений.
Концентрация водородных ионов (рН) нередко сказывается на распределении водных организмов. Пресноводные бассейны с рН 3,7— 4,7 считаются кислыми, 6,95—7,3 нейтральными, с рН более 7,8 — щелочными. В пресных водоемах рН испытывает значительные колебания, нередко в течение суток. Морская вода более щелочная, и рН ее меньше изменяется, чем пресной. С глубиной рН уменьшается.
Из растений при рН меньше 7,5 растут полушник (Jsoetes), ежеголовник (Sparganium). В щелочной среде (рН 7,7—8,8) распространены многие виды рдестов, элодея, при рН 8,4—9 сильного развития достигает Typha angustifolia. Кислые воды торфяников способствуют развитию сфагновых мхов.
Большинство пресноводных рыб выдерживает рН от 5 до 9. Если рН меньше 5, наблюдается массовая гибель рыб, а выше 10 — погибают все рыбы и другие животные.
В озерах с кислой средой часто встречаются личинки двукрылых из рода Chaoborus, а в кислых водах болот распространены раковинные корненожки (Testaceae), отсутствуют пластинчато-жаберные моллюски из рода беззубок (Unio), редко встречаются другие моллюски.
Экологическая пластичность организмов водной среды. Вода является более стабильной средой, и абиотические факторы претерпевают сравнительно незначительные колебания, а отсюда водные организмы обладают по сравнению с наземными меньшей экологической пластичностью. Пресноводные растения и животные более пластичны, чем морские, так как пресная вода как среда жизни более изменчива. Оценивают широту экологической пластичности гидробионтов не только в целом к комплексу факторов (эври- и стенобионтность), но и по отдельности.
Так, установлено, что прибрежные растения и животные в отличие от обитателей открытых зон главным образом эвритермные и эвригалинные организмы, вследствие того, что температурные условия и солевой режим вблизи берега довольно изменчивы — прогреваемость солнцем и относительно интенсивное охлаждение, опреснение притоком воды из ручьев и рек, в частности в период дождей, и т. д. В качестве примера можно привести лотос, который относится к типичным стенотермным видам, произрастает только в мелких, хорошо прогреваемых водоемах. Обитатели поверхностных слоев по сравнению с глубоководными формами по вышеуказанным причинам оказываются более эвритермными и эвригалинными.
Экологическая пластичность является важным регулятором расселения организмов. Доказано, что гидробионты с высокой экологической пластичностью распространены широко, например, элодея. Противоположный же пример — рачок артемия (Artemia solina), живущий в небольших водоемах с очень соленой водой, является типичным стеногалинным представителем с узкой экологической пластичностью. По отношению же к другим факторам он обладает значительной пластичностью и в соленых водоемах встречается довольно часто.
Экологическая пластичность зависит от возраста и фазы развития организма. Например, морской брюхоногий моллюск Littorina во взрослом состоянии при отливах ежедневно длительное время обходится без воды, однако его личинки ведут планктонный образ жизни и не переносят высыхания.
Особенности адаптации растений к водной среде. Водные рай| стения имеют значительные отличия от наземных растительных организмов. Так, способность водных растений поглощать влаг и минеральные соли непосредственно из окружающей среди отражается на их морфологической и физиологической органй зации. Характерным для водных растений является слабое развив тие проводящей ткани и корневой системы. Корневая системой служит главным образом для прикрепления к подводному субЙ страту и не выполняет функции минерального питания и водоснабжения, как у наземных растений. Питание же водных растений осуществляется всей поверхностью их тела.
Значительная плотность воды дает возможность обитания растений во всей ее толще. У низших растений, заселяющих различные слои и ведущих плавающий образ жизни, для этого имеются специальные придатки, которые увеличивают их плавучесть позволяют им удерживаться во взвешенном состоянии. Высшй гидрофиты имеют слабо развитую механическую ткань. Как yni отмечалось выше, в их листьях, стеблях, корнях располагаютс воздухоносные межклеточные полости, увеличивающие легкость и плавучесть взвешенных в воде и плавающих на поверхност органов, что также способствует смыванию внутренних клетой водой с растворенными в ней солями и газами. Гидрофиты отли| чаются большой поверхностью листьев при малом общем объеме растения, что обеспечивает им интенсивный газообмен при недостатке растворенного в воде кислорода и других газов.
У ряда водных организмов развита разнолистность, или гетё рофилия. Так, у сальвинии (Salvinia) погруженные листья обеспечивают минеральное питание, а плавающие — органическое.
Важной особенностью адаптации растений к обитанию в вод| ной среде является и то, что листья, погруженные в воду, как правило, очень тонкие. Часто хлорофилл в них располагается в клетках эпидермиса, что способствует усилению интенсивности фотосинтеза при слабом освещении. Такие анатомо-морфологические особенности наиболее четко выражены у водяных мхов (Riccia, Fontinalis), валиснерии (Vallisneria spiralis), рдестов (Potamageton).
От вымывания у водных растений из клеток минеральных солей или выщелачивания защитой является выделение специальными клетками слизи и образование эндодермы из более толстостенных клеток в виде кольца.
Относительно низкая температура водной среды обусловливает отмирание вегетирующих частей у погруженных в воду растений после образования зимних почек и замену летних тонких нежных листьев более жесткими и короткими зимними. Низкая температура воды отрицательно сказывается на генеративных органах водных растений, а высокая ее плотность затрудняет перенос пыльцы. В связи с этим водные растения интенсивно размножаются вегетативным путем. Большинство плавающих на поверхности и погруженных растений выносят цветоносные стебли в воздушную среду и размножаются половым путем. Пыльца разносится ветром и поверхностными течениями. Плоды и семена, которые образуются, также распространяются поверхностными течениями. Это явление носит название гидрохории. К гидрохорным относятся не только водные, а также многие прибрежные растения. Их плоды имеют высокую плавучесть, длительное время находятся в воде и не теряют при этом всхожесть. Например, водой переносятся плоды и семена стрелолиста (Sagittaria sagittofolia), сусака (Butomus umbellatus), частухи (Alisma plantago-aguatica). Плоды многих осок (Carex) заключены в своеобразные мешочки с воздухом и разносятся водными течениями. Таким же образом расселился сорняк гумай (Sorgnum halepense) вдоль реки Вахт по каналам.
Особенности адаптации животных к водной среде. У животных, обитающих в водной среде, по сравнению с растениями адаптивные особенности более многообразны, к ним относятся такие, как анатомо-морфологические, поведенческие и др.
Животные, обитающие в толще воды, обладают в первую очередь приспособлениями, которые увеличивают их плавучесть и позволяют противостоять движению воды, течениям. Данные же организмы вырабатывают приспособления, которые препятствуют поднятию их в толщу воды или уменьшают плавучесть, что позволяет удержаться на дне, включая и быстро текущие воды.
У мелких форм, живущих в толще воды, отмечается редукция скелетных образований. Так, у простейших (Radiolaria, Rhizopoda) раковины обладают пористостью, кремневые иглы скелета внутри полые. Удельная плотность гребневиков (Ctenophora), медуз (Scyphozoa) уменьшается благодаря наличию воды в тканях. Скопление капелек жира в теле (ночесветки — Noctiluca, радиолярии — Radiolaria) способствует увеличению плавучести. Крупные скопления жира наблюдаются у некоторых ракообразных (Cladocera, Copepoda), рыб и китообразных. Удельную плотность тела снижают и тем самым повышают плавучесть плавательные пузыри, наполненные газом, которые имеют многие рыбы. У сифонофор (Physalia, Velella) развиты мощные воздухоносные полости.
Для животных, пассивно плавающих в толще воды, характерно не только уменьшение массы, но и увеличение удельной поверхности тела. Это связано с тем, что чем больше вязкость среды и выше удельная поверхность тела организма, тем он медленнее погружается в воду. У животных уплощается тело, на нем образуются шипы, выросты, придатки, например у жгутиковых (Leptodiscus, Craspeditella), радиолярий (Aulacantha, Chalengeridae) и др.
Большая группа животных, обитающих в пресной воде, при передвижении использует поверхностное натяжение воды (поверхностную пленку). По поверхности воды свободно бегают клопы водомерки (Gyronidae, Veliidae), жуки вертячки (Gerridae) и др. Членистоногое, касающееся воды окончанием своих придатков, покрытых водоотталкивающими волосками, вызывает деформацию ее поверхности с образованием вогнутого мениска. Когда подъемная сила (F), направленная вверх, больше массы животного, последнее и будет удерживаться на воде благодаря поверхностному натяжению.
Таким образом, жизнь на поверхности воды возможна для сравнительно мелких животных, так как масса растет пропорционально кубу размера, а поверхностное натяжение увеличивается как линейная величина.
Активное плавание у животных осуществляется с помощью ресничек, жгутиков, изгибания тела, реактивным способом за счет энергии выбрасываемой струи воды. Наибольшего совершенства реактивный способ передвижения достигу головоногих моллюсков. Так, некоторые кальмары развивают скорость при выбрасывании воды до 40—50 км/ч (рис. 5.12).

Рис. 5.12. Кальмар
У крупных животных нередко имеются специализированные конечности (плавники, ласты), тело их обтекаемой формы и покрыто слизью.
Только в водной среде встречаются неподвижные, ведущие прикрепленный образ жизни, животные. Это такие, как гидроиды (Hydroidea) и коралловые полипы (Anthozoo), морские лилии (Crinoidea), двустворчатые (Вг/аМа)идр.Дляних характерны своеобразная форма тела, незначительная плавучесть (плотность тела больше плотности воды) и специальные приспособления для прикрепления к субстрату.
Водные животные большей частью пойкилотермны. У гомой-отермных же, например, млекопитающих (китообразные, ластоногие) образуется значительный слой подкожного жира, который выполняет теплоизоляционную функцию.
Глубоководные животные отличаются специфическими чертами организации: исчезновение или слабое развитие известкового скелета, увеличение размеров тела, нередко — редукция органов зрения, усиление развития осязательных рецепторов и т. д.
Осмотическое давление и ионное состояние растворов в теле животных обеспечивается сложными механизмами водно-солевого обмена. Наиболее распространенным способом поддержания постоянного осмотического давления является регулярное удаление поступающей в организм воды с помощью пульсирующих вакуолей и органов выделения. Так, пресноводные рыбы избыток воды удаляют усиленной работой выделительной системы, а соли поглощают через жаберные лепестки. Морские же рыбы вынуждены пополнять запасы воды и поэтому пьют морскую воду, а излишки поступающих с водой солей выводят из организма через жаберные лепестки (рис. 5.13).
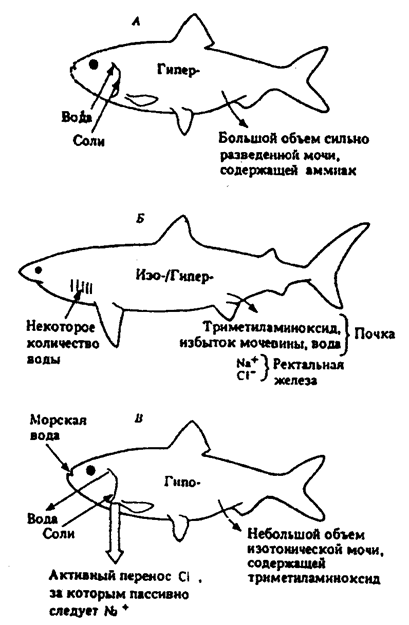
Рис. 5.13. Экскреция и осморегуляция у пресноводных костистых
рыб (А), пластинчатожаберных (Б) и морских костистых рыб (В)
Сокращения гипо-, изо- и гипер- указывают тоничность внутренней среды по отношению к внешней (из Н. Грина и др., 1993)
Целый ряд гидробионтов обладают особым характером питания — это отцеживание или осаждение взвешенных в воде частиц органического происхождения, многочисленных мелких организмов. Этот способ питания не требует больших затрат энергии на поиски добычи и характерен для пластинчатожаберных моллюсков, сидячих иглокожих, асцидий, планктонных рачков и др. Животные-фильтраторы выполняют важную роль в биологической очистке водоемов.
Пресноводные дафнии, циклопы, а также самый массовый в океане рачок Calanus finmarchicus отфильтровывают в сутки до 1,5 л воды на особь. Мидии, обитающие на площади 1 м2, могут прогонять через мантийную полость 150—280 м3 воды за сутки, осаждая взвешенные частицы.
В связи с быстрым затуханием световых лучей в воде жизнь в постоянных сумерках или во мраке сильно ограничивает возможности зрительной ориентации гидробионтов. Звук распространяется в воде быстрее, чем в воздухе, и ориентация на звук у гидробионтов развита лучше зрительной. Отдельные виды улавливают даже инфразвуки. Звуковая сигнализация служит больше всего для внутривидовых взаимоотношений: ориентации в стае, привлечения особей другого пола и т. д. Китообразные, например, отыскивают пищу и ориентируются при помощи эхолокации — восприятия отраженных звуковых волн. Принцип локатора дельфина заключается в излучении звуковых волн, которые распространяются перед плывущим животным. Встречая препятствие, например рыбу, звуковые волны отражаются и возвращаются к дельфину, который слышит возникающее эхо и таким образом обнаруживает предмет, вызывающий отражение звука.
Известно около 300 видов рыб, которые способны генерировать электричество и использовать его для ориентации и сигнализации. Ряд рыб (электрический скат, электрический угорь и др.) используют электрические поля для защиты и нападения.
Водным организмам свойствен древний способ ориентации — восприятие химизма среды. Хеморецепторы многих гидробионтов (лососи, угри и др.) обладают чрезвычайной чувствительностью. В тысячекилометровых миграциях они с поразительной точностью находят места нерестилищ и нагула.
Смена условий в водной среде вызывает и определенные поведенческие реакции организмов. С изменением освещенности, температуры, солености, газового режима и других факторов связаны вертикальные (опускание вглубь, поднятие к поверхности) и горизонтальные (нерестовые, зимовальные и нагульные) миграции животных. В морях и океанах в вертикальных миграциях принимают участие миллионы тонн гидробионтов, а при горизонтальных миграциях водные животные могут преодолевать сотни и тысячи километров.
На Земле существует много временных, неглубоких водоемов, возникающих после разлива рек, сильных дождей, таяния снега и т. д. Общими особенностями обитателей пересыхающих водоемов является способность давать за короткие сроки многочисленное потомство и переносить длительные периоды без воды, переходя в состояние пониженной жизнедеятельности —гипобиоза.
Дата добавления: 2015-02-13; просмотров: 1810;