Характеристика пресноводных экосистем
Лентические экосистемы в литоральной зоне содержат два типа продуцентов: укрепившиеся в дне цветковые растения и плавающие зеленые растения ¾ водоросли, некоторые высшие (рдесты) (рис. 7.7). Растения, укрепленные в дне, образуют три концентрические зоны:
1) зона надводной вегетации ¾ фотосинтезирующая часть растений (камыши, рогозы и др.) находится над водой, а биогенные элементы извлекаются из донных осадков;
2) зона укрепленных в дне растений с плавающими по воде листьями (кувшинки) ¾ у них та же роль, что и у растений первой зоны, но они могут затенять нижние толщи воды;
3) зона подводной вегетации ¾ укорененные и прикрепленные растения полностью находящиеся под водой и осуществляющие фотосинтез и минеральный обмен в водной среде (рдесты и прикрепленные водоросли ¾ харовые).

Рис. 7.7. Некоторые продуценты лентических сообществ; прибрежные растения,
укореняющиеся в дне водоема (1-7), нитчатые водоросли (8-9)
и фитопланктон (10-20)
Животные, консументы, более разнообразны в литорали, чем в других зонах водоема. Перифитон представлен моллюсками, коловратками, мшанками, личинками насекомых и др. Многие животные нектона дышат кислородом атмосферного воздуха (лягушки, саламандры, черепахи и др.). Рыбы большую часть жизни проводят в литорали и здесь же размножаются. Зоопланктон представлен ракобразными, имеющими большое значение для питания рыб (дафнии и др.).
В сообществах лимнической зоны продуцентом является фитопланктон. В водоемах умеренного пояса плотность его популяции заметно изменяется по сезонам. Весной «цветение» связано с массовым развитием приспособленных к прохладной воде диатомитовых водорослей, летом ¾ зеленых, осенью ¾ азотфиксирующих синезеленых водорослей. Зоопланктон представлен растительноядными ракоообразными и коловратками, все другие ¾ хищники. Нектон лимнической зоны ¾ это только рыбы.
Сообщества профундальной зоны существуют без света. Фауна и флора здесь ¾ в зоне поверхностного раздела вода ¾ ил, где накапливается органический материал, ¾ представлена бактериями и грибами (редуценты), а также бентосными формами ¾ личинками насекомых, моллюсками, кольчатыми червями (консументами).
Количество красных кольчатых червей возрастает с ростом загрязнения водоема сточными водами, т. е. по этому показателю можно судить о степени загрязнения водоема.
Действие на сообщества стоячих водоемов таких лимитирующих факторов, как содержание кислорода, температуры и освещенности, зависит от специфических особенностей этих водоемов ¾ озер, прудов, искусственных водохранилищ.
Озера¾ естественные пресноводные водоемы (см. рис. 7.9), образовались геологически сравнительно недавно ¾ за последние несколько десятков тысяч лет, и лишь возраст некоторых из них исчисляется миллионами лет, например, Байкала. Наличие у большинства озер профундальной зоны сказывается на температурном режиме водной толщи, на ее «перемешивании» и распределении кислорода в ней. Эти процессы сезонны, как и стратификация озера по температурному режиму (рис. 7.8).
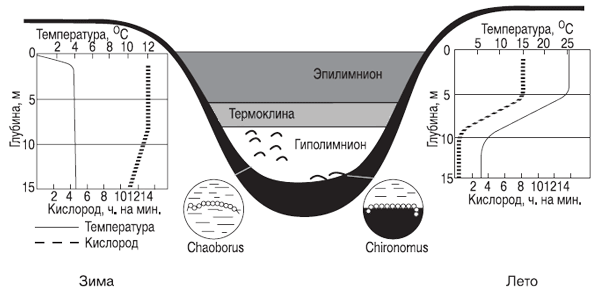
Рис. 7.8. Температурная стратификация в озере умеренной зоны Северного полушария
(оз. Линсли, Коннектикут, США, по Ю. Одуму, 1975)
В озерах умеренного пояса в летнее время можно выделить в вертикальном разрезе три зоны: эпилимнион ¾ до глубины, где происходит конвекция (циркуляция) воды; термоклина ¾ это промежуточная зона, где вода не смешивается с водой верхней зоны; гиполимнион ¾ область холодной воды, где нет циркуляции.
Термоклина обычно расположена ниже границы проникновения света, и запасы кислорода, в отрезанном от его источников гиполимнионе, истощаются. Наступает летний период стагнации. Осенью, вследствие выравнивания температур, происходит общее перемешивание воды и обогащение гиполимниона кислородом. Зимой, когда температура воды под льдом становится ниже плюс 4 °С, что снижает ее плотность и снова приводит к стратификации озера и к зимней стагнации. Весной, после таяния льда, температура воды достигает плюс 4 °С, она тяжелеет и снова происходит весеннее перемешивание. Это классическая схема для водоемов Евразии и Северной Америки. В полярных областях и субтропиках перемешивание бывает только один раз в году: в первом случае ¾ летом, во втором ¾ зимой. В водоемах тропиков перемешивание воды идет постоянно, но медленно, а общее перемешивание происходит редко и нерегулярно.
Цветение фитопланктона обычно приурочено к перемешиванию, когда в эвфотической зоне появляются воды, обогащенные природными биогенными компонентами. С точки зрения продуктивности озера подразделяются на две группы: 1) олиготрофные (малокормные) и 2) эвтрофные (кормные). Продуктивность лентических экосистем зависит также от поступающих веществ с окружающей суши и от глубины озера (наиболее продуктивны мелкие озера).
Пруды обладают хорошо развитой литоралью и практическим отсутствием стратификации, образуются они в различных понижениях, часто временно пересыхают летом или в засушливые годы. Фауна прудов способна переживать сухие периоды в покоящемся состоянии или перебираться в другие водоемы (земноводные). Естественные пруды ¾ высокопродуктивны. В искусственных прудах, в основном, человек сам подкармливает рыб.
Водохранилища (см. рис. 7.9) создаются человеком при возведении гидроэнергетических и гидромелиоративных комплексов. Это уже не природная экосистема, а природно-техническая система. Распределение тепла и биогенов в ней зависит от типа плотины. Если вода сбрасывается придонная, то в этом случае водохранилище аккумулирует тепло и экспортирует биогенные вещества, если сброс идет поверх плотины, то экспортируется тепло и аккумулируются биогены. В первом случае спускается вода гиполимниона, во втором ¾ эпилимниона. Через глубоководные шлюзы в реку поступает и более соленая вода, а биогены вызывают эвтрофикацию участка реки.

Рис. 7.9. Пресноводные экосистемы
Лотические экосистемы¾реки (см. рис. 7.9) отличаются от стоячих водоемов тремя условиями:
1) течение ¾ важный лимитирующий и контролирующий фактор;
2) обмен между водой и сушей значительно более активен;
3) распределение кислорода более равномерно, так как практически отсутствует стратификация.
Скорость течения влияет на распределение рыб в реках ¾ они могут жить и под камнями, и в заводях, под перекатами, но это будут разные виды, адаптированные к конкретным условиям. Река ¾ открытая экосистема, в которую поступает с прилегающих пространств большое количество органического вещества.
Детритное питание ¾ основа трофических цепей лотических экосистем: более 60% энергии консументы получают от привнесенного материала. Зато кислорода в реках достаточно и содержание его в воде постоянно, что обусловило узкую толерантность организмов по отношению к кислороду.
Выделяют лотические сообщества перекатов и плесов. На перекатах поселяются организмы, способные прикрепиться к субстрату (нитчатые водоросли), или хорошие пловцы (форель). На участках плеса сообщества напоминают прудовые.
В больших реках прослеживается продольная зональность: в верховьях ¾ сообщества перекатов, в низовьях и дельте ¾ плесов, между ними местами могут возникать и те и другие. Продольная зональность подчеркивается изменениями видового состава рыб. К низовьям видовой состав обедняется, но увеличиваются размеры рыб.
Заболоченные пресноводные участки, обычно¾собственно болота (см. рис. 7.9) ¾ низинные и верховые. Низинные имеют, как правило, питание подземными водами, а верховые ¾ атмосферными осадками. Верховые могут встречаться в любом понижении или даже на склонах гор, низинные возникают вследствие зарастания озер и речных стариц. Они покрыты водными макрофитами, болотными растениями и кустарниками.
Болотные почвы и торфяники содержат много углерода (14-20%), сельскохозяйственная отработка которых приводит к выделению в атмосферу большого количества углекислого газа, что усугубляет «CO2-проблему».
Дата добавления: 2015-02-10; просмотров: 2637;