Репликация
В процессе копирования информации происходит синтез дочерних молекул ДНК на основе информации, "записанной" в родительской молекуле ДНК. Ясно, что дочерние молекулы должны представлять собой точные копии родительской.
Репликация может осуществляться тремя способами:
а) консервативным;
б) полуконсервативным;
в) дисперсивным.
При консервативной репликации вновь синтезированные цепи ДНК находятся в дочерней молекуле. При полуконсервативной репликации полученные молекулы состоят из родительской и вновь синтезированной цепей. Дисперсивный способ репликации означает наличие перемежающихся родительских и вновь синтезированных участков на каждой из цепей образованных молекул ДНК. Для животных организмов и человека характерен только полуконсервативный путь репликации ДНК.
В процессе репликации участвует целый ряд энзимов (ферментов) с определенными функциями. Только синтезирующих ферментов в клетках прокариот насчитывается три. Их называют ДНК - полимеразами I, II и III. Сведения о функциональных особенностях ДНК-полимераз приведены в таблице.
| Проявляемая функция (активность) | Полимераза | |||
| Pol I | Pol II | Pol III | Фрагмент Кленова | |
| Полимеразная 5'-3' | есть | есть | есть | есть |
| Экзонуклеазная 3'-5' | есть | есть | есть | есть |
| Экзонуклеазная 5'-3' | есть | нет | нет | нет |
| молекул в клетке | нет данных | 10-20 | нет | |
| производительность (нуклеотидов в мин, 37 С, на 1 молекулу Pol) | - |
Фрагмент Кленова – результат частичного протеолиза ДНК-полимеразы I E. Coli субтилизином. Основная функция полимеразы III – синтез цепи, полимеразы I – синтез и исправление ошибочно вставленных нуклеотидов. Полимераза II осуществляет особые, специализированные функции.
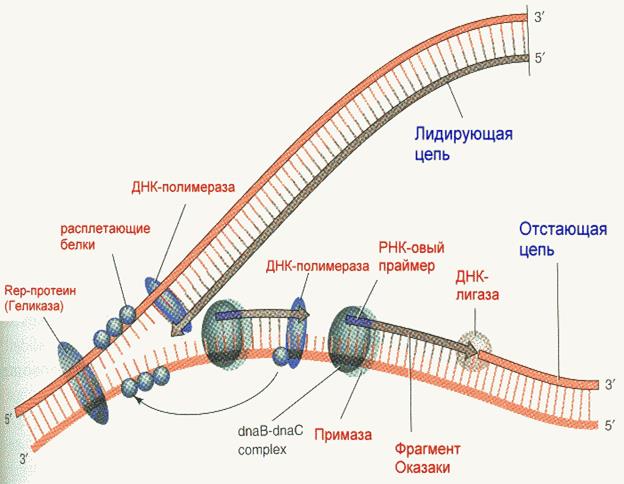
Репликация начинается с расплетания цепей ДНК специальными расплетающими белками, которые называют геликазами (или Rep-протеином). Геликазы используют энергию АТФ в процессе расплетания цепей. Скорость расплетания составляет около 6000 мин-1. Для того чтобы расплетенные цепи не могли вновь соединиться, имеются специальные SSB-белки (single-strand binding proteins), которые присоединяются к комплементарным цепям, удерживая их от ассоциации. По мере продвижения репликационной вилки SSB-протеины передвигаются по цепи, диссоциируя с одного места и присоединяясь на другом. Этот процесс не требует затрат энергии АТФ. После освобождения достаточного места начинается синтез праймера - затравки, необходимой для работы ДНК-полимеразы. Наличие затравки является необходимым условием функционирования ДНК-полимераз (как и наличие комплементарной цепи). В качестве затравки на каждой из разделенных цепей синтезируются маленькие отрезки молекул РНК при помощи фермента примазы.
Синтез новой цепи ДНК осуществляется всегда в направлении 5'-3', поэтому если по одной матричной цепи возможен непрерывный синтез, то по комплементарной ей цепи синтез осуществляется только участками. Эти участки синтеза называют фрагментами Оказаки. Когда синтез на одном из фрагментов Оказаки достигает праймера другого фрагмента, РНК-овый праймер удаляется имеющейся у полимераз 5'-3' полимеразной активностью и достраивается дезоксирибонуклеотидами. После этого сахарофосфатный остов между фрагментами сшивается ковалентной связью при помощи фермента ДНК-лигазы.
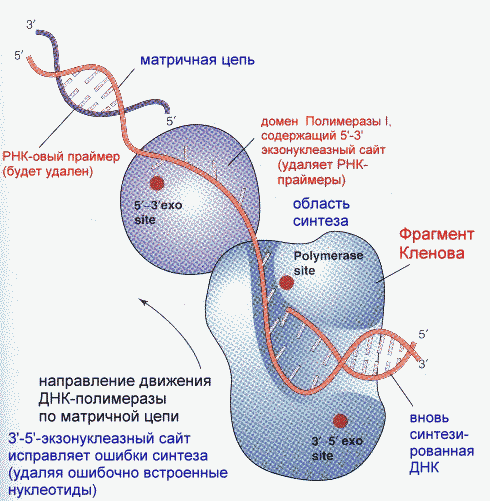
Частота возникновения ошибок при репликации и транскрипции не превышает 10-8-10-9, то есть, возможна лишь одна ошибка на сотни миллионов нуклеотидов. Такая точность не может быть обеспечена одним только лишь правилом комплементарности нуклеотидов (обеспечивающим точность 1:10000-1:100000). Репликационный аппарат имеет собственные механизмы "поддержания точности" копирования генетической информации. Этими функциями наделена ДНК-полимераза I.
Модель ее структуры и функциональных участков показана на рисунке. Она имеет три зоны активности: полимеризующую в направлении 5'-3', и экозонуклеазные в направлениях 5'-3' и 3'-5'. Области активности разделены пространственно. Вперед (по ходу продвижения полимеразы по матричной цепи ДНК) обращена зона 5'-3' экзонуклеазной активности. Она служит для удаления попадающихся на пути РНК-овых праймеров (затравок). Далее идет собственно синтетическая зона и, наконец, зона с экзонуклеазной активностью в направлении 3'-5'. С этой зоной связана так называемая PROOF-READING активность (способность узнавать неправильно встроенные нуклеотиды) и исправлять их вырезанием ряда уже встроенных нуклеотидов. Для этого молекула ДНК-полимеразы смещается (не отсоединяясь от ДНК-овой матрицы) к месту синтеза и последовательно вырезает нуклеотиды, после чего возобновляется нормальный синтез.
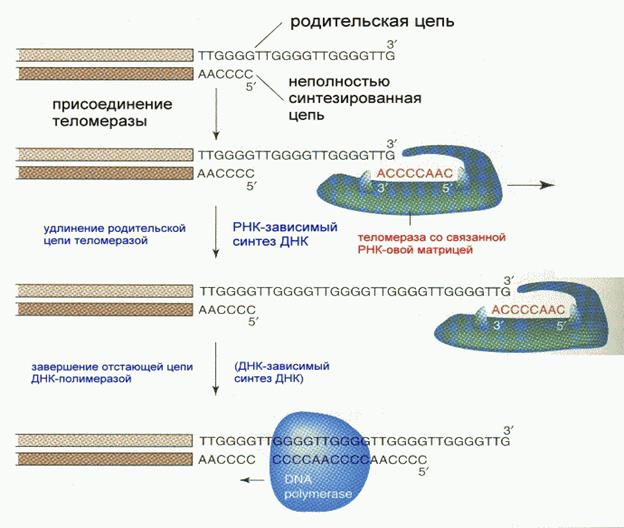
В процессе репликации одна из вновь синтезированных цепей (а именно- синтезированная на лидирующей цепи) получается короче на несколько десятков нуклеотидов вследствие того, что содержала РНК-овый праймер (затравку), впоследствии удаленный. Однако получение более коротких копий является совершенно недопустимым в процессе репликации явлением. Положение исправляется при помощи так называемых теломераз. Теломераза, содержащая в себе последовательность нуклеотидов, за несколько приемов удлиняет укороченную цепь, создавая пространство для работы примазы и ДНК-полимеразы, после чего избытки нуклеотидов удаляются:
Воздействие на организм неблагоприятных факторов (химические соединения, ультрафиолет и др.) приводит к постоянному накоплению ошибок в геноме, которые, в конечном итоге, вызывают появление патологии, в частности, невыясненный до сих пор механизм раковых заболеваний. Пока лишь существуют только предположения о том, что причиной раковых заболеваний являются дефекты в носителях информации- ДНК.
Дата добавления: 2015-02-07; просмотров: 1587;