Ближний и дальний транспорт ионов в растениях
В зависимости от уровня организации процесса различают три типа транспорта веществ в растении: внутриклеточный, ближний (внутри органа) и дальний (между органами).
Внутриклеточный транспорт. Передвижение веществ внутри одной клетки осуществляется в результате совместного действия циклозиса (круговое движение цитоплазмы) и направленной поперек этого движения диффузии, чем может достигаться почти полное перемешивание веществ в гиалоплазме. У высших растений движение цитоплазмы происходит при участии сократительных белков актомиозинового типа. Скорость движения цитоплазмы 0,2-0,6 мм/мин. Во внутриклеточном транспорте веществ принимают участие также каналы эндоплазматического ретикулума и везикулы Гольджи.
Ближний транспорт. Это передвижение ионов, метаболитов и воды между клетками и тканями внутри органа. Ближний транспорт включает радиальный транспорт веществ в корнях и стеблях, передвижение веществ в мезофилле листьев на небольшие расстояния, измеряемые миллиметрами. Осуществляется он через клетки неспециализированных для транспорта веществ тканей по апопласту - совокупности межклетников и межфибриллярных полостей клеточных стенок, симпласту - совокупности протопластов клеток, соединенных плазмодесмами и вакуому - дискретной системе вакуолей клеток.
Дальний транспорт. Это передвижение веществ между органами растения. Осуществляется по специализированной проводящей системе, включающей сосуды и трахеиды ксилемы (восходящий ток) и ситовидные трубки флоэмы (нисходящий ток).
Радиальное перемещение ионов в корне. Путем обменных процессов и диффузии ионы поступают в клеточные стенки ризодермиса, а затем через коровую паренхиму направляются к проводящим пучкам. Вплоть до внутреннего слоя коры эндодермы возможно перемещение ионов по апопласту за счет диффузии и обменной адсорбции. Транспирационный ток воды ускоряет этот процесс. Поглощение ионов клетками коры может происходить на всем пути их движения по апопласту, что увеличивает фактическую поверхность соприкосновения корня с внешним раствором.
Однако в обычных условиях существования, при сравнительно низких концентрациях ионов в наружной среде, их первичное поглощение осуществляется главным образом ризодермисом. Основным местом входа ионов в симпласт являются корневые волоски. Одним из косвенных доказательств этого служит высокая насыщенность плазмодесмами базальной стенки трихобласта на его границе с корой. Ризодермис с корневыми волосками функционирует как нагнетательный насос. Именно здесь происходит загрузка ионами корневого симпласта, а далее вплоть до сосудов они проходят весь путь по плазмодесмам внутри симпластической фазы без преодоления мембранных барьеров.
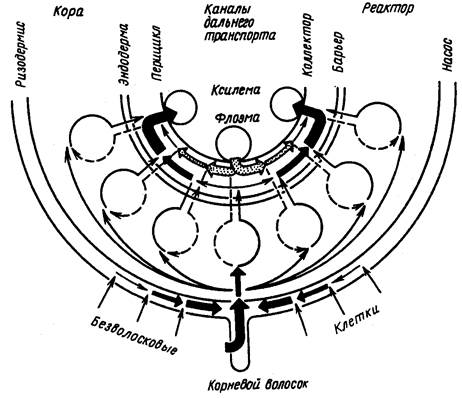
Симпластический транспорт веществ сопровождается их метаболизацией, т.е. включением в процессы обмена веществ. Поэтому кору называют метаболическим реактором корня. Здесь минеральные формы азота, фосфора и серы включаются в органические соединения и к сосудам уже направляются продукты их первичной ассимиляции. Кора выполняет также функцию транспортного буфера, регулирующего ионный поток по радиусу корня. При высоком уровне питания избыток ионов изымается из симпластического потока путем сбрасывания их в вакуоли и к сосудам подается необходимое количество питательных веществ.
При дефицитном питании ионные потоки на тонопласте смещаются в сторону симпласта и недостаток ионов смягчается запасами вакуоли. При отсутствии ионов в среде, например, при помещении корней в воду в условиях эксперимента наблюдается «тканевый плач» с достаточно высокой концентрацией ионов в пасоке в основном за счет содержимого вакуолей коры, которые составляют до 80 % общего вакуолярного объема корня.
Буферная роль корня наглядно проявляется в существенном изменении содержания азота, фосфора, калия и других питательных элементов в корне и относительном постоянстве их содержания в побеге при изменении условий питания. Так, при изменении обеспеченности фосфором в 4 раза (выращивание на 1/2 и двух дозах питательной смеси) его содержание в корнях увеличивалось в 3 раза, а в листьях практически не изменялось.
Это надо иметь в виду при листовой диагностике. Более объективную информацию по обеспеченности сельскохозяйственных культур элементами минерального питания дает не содержание элементов в листьях, а соотношение их содержания в листьях и корнях, называемое коэффициентом распределения.
Особое значение в радиальном транспорте ионов имеет самый внутренний слой клеток коры - эндодерма. Пояски Каспари прерывают апопластический транспорт веществ между косой и центральным цилиндром. Весь поток переходит на симпластический путь, чем обеспечивается метаболический контроль поступления веществ. Кроме того, эндодерма препятствует утечке ксилемного сока из центрального цилиндра. Следующая ткань - перицикл работает как кольцевой коллектор, направляя вещества кратчайшим путем из системы симпласта к сосудам ксилемы.
Механизмы преодоления первой мембраны симпласта и его последней мембраны различны. Если загрузка симпласта в ризодермисе происходит активно с участием Н-помпы, то его разгрузка (около сосудов) происходит, вероятно, пассивно. Имеются экспериментальные данные, позволяющие считать, что функционирование Н-помпы в мембранах околососудистых клеток обеспечивает не выделение К в сосуды, а, наоборот, его обратное поглощение из сосудов клетками паренхимы центрального цилиндра, что обусловливает рециркуляцию ионов и стабилизацию ионного состава ксилемного сока.
Восходящий ток ионов осуществляется преимущественно по сосудам ксилемы, которые лишены живого содержимого и являются составной частью апопласта растения. Механизм ксилемного транспорта - массовый ток растворенных веществ вместе с водой, движимый транспирацией и корневым давлением. Но и на этом этапе транспорта веществ, так же как и в корне, осуществляется его активная регуляция.
Применение радиоактивных изотопов фосфора и калия позволило обнаружить, что, хотя они и движутся вместе с восходящим транспирационным током, тем не менее в большем количестве они поступают не в более энергично транспирирующие, а в растущие части растения - в точки роста, только развертывающиеся листочки, в формирующиеся плоды. Движение ионов по сосудам ксилемы происходит при их непрерывном взаимодействии со стенками самих сосудов. В клеточных стенках ксилемы имеются многочисленные участки с фиксированными отрицательными зарядами, способные избирательно связывать катионы и таким образом тормозить их транспорт по ксилеме.
Ионный поток в ксилеме активно регулируется также прилегающими живыми клетками. Если какого-то элемента в ксилемном соке слишком много, он извлекается из сосудов. Они же могут служить и источником недостающих элементов питания, Предавая их по мере необходимости в ксилемный сок. Таким образом, стебель представляет вторую после корня буферную емкость, обеспечивающую равномерность поступления веществ и ионный гомеостаз растительного организма. Благодаря этому в растении при продвижении растущих корней через почвенные слои, в разной мере обеспеченные питательными веществами нет перебоев в снабжении ими надземных органов.
Использование метода меченых атомов (32Р, 42K.) позволило выявить участие в восходящем токе веществ элементов коры. Ее вклад варьирует в зависимости от обеспеченности растении минеральными веществами и водой. Такое дублирование повышает надежность транспортной системы. Пространственная сближенность ксилемы с сосудами флоэмы структурно обеспечивает интенсивный диффузионный обмен между ними уже в стебле. Здесь значительная часть калия вовлекается в обратный базинетальный транспорт, не достигая листьев. Такая рециркуляция калия как осмотически активного вещества может существенно повышать количество воды, транспортируемой по ксилеме.
Состав ксилемного сока. Ксилемный сок представляет собой раствор, в основном состоящий из неорганических веществ. Однако в пасоке, вытекающей из ксилемы пенька при удалении верхней части стебля, обнаружены также различные азотистые соединения (аминокислоты, амиды, алкалоиды и др.), органические кислоты, фосфорорганические эфиры, соединения, содержащие серу, некоторое количество сахаров и многоатомных спиртов, а также фитогормоны. В ксилемном соке могут содержаться и более сложные вещества, попадающие сюда из вакуолей и цитоплазмы трахеальных элементов, заканчивающих свое развитие.
Сравнительный состав (в ммоль/л) флоэмного и ксилемного сока у однолетнего люпина
| Вещества | Ксилема | Флоэма |
| Сахароза | - | 450 - 470 |
| Аминокислоты, в мг/мл | 0,7 - 2,6 | 13 - 15 |
| Калий | 2,4 - 4,6 | 39,0 - 46,0 |
| Натрий | 2,2 - 2,6 | 4,4 - 5,2 |
| Кальций | 0,4 - 1,8 | 0,5 - 1,6 |
| Магний | 0,3 - 1,1 | 3,5 - 5,8 |
| Нитраты | Следы | 0,003 |
| рН | 5,9 | 8,0 |
Органические компоненты ксилемного сока меняются в зависимости от вида растения и от природы ионов, присутствующих в наружном растворе. Если поместить корни в солевой раствор, из которого катионы поглощаются легче, чем анионы, то количество поглощенных катионов превысит количество анионов. Растение компенсирует эту несбалансированность синтезом органических анионов, обычно карбоновых кислот: сукцината, малата, цитрата и др. В условиях, обеспечивающих высокую нитратредуктазную активность корней, важным компонентом ксилемного сока могут быть аминокислоты глутамат, аспартат, лизин и др.
Ксилемный сок по составу резко отличается от вакуолярного. Например, содержание иона К+ в вакуолях эпикотиля гороха достигает 55 - 78 ммоль/л, а в ксилемном соке - лишь 2-4 ммоль/л.
Поглощение ионов клетками листьев. На долю проводящей системы приходится около 1/4 объема ткани листа. Суммарная длина разветвлений проводящих пучков в 1 см2 листовой пластинки достигает 1 м. Такая насыщенность тканей листа проводящей системой обеспечивает ее тесный контакт с клетками мезофилла и разгрузку ксилемы в листе. Вода и растворенные в ней вещества через поры сосудов ксилемы попадают как в клеточные стенки (апопласт), так и в цитоплазму клеток мезофилла. Но диффузия ионов по свободному пространству листа затруднена из-за крайне сложной его формы. Поэтому большая часть ионов поступает внутрь клеток мезофилла при участии Н+ помпы.
Непрерывная доставка ионов в клетки мезофилла обеспечивает растущий лист неорганическими солями и азотистыми соединениями, необходимыми для новообразования веществ и поддержания осмотического потенциала растительных тканей. В зрелом листе, достигшем конечных размеров, поступление веществ с транспирационным током превосходит потребности в них и есть опасность значительного повышения осмотического давления. В тканях листьев используются три способа устранения избытка ионов: образование труднорастворимых осадков солей (карбонатов кальция в клеточных стенках, оксалата кальция в вакуолях, фосфата кальция в митохондриях); отток ионов из листа через флоэму; накопление и выделение солей солевыми желёзками и волосками.
Большинству культур, произрастающих на незасоленных почвах, свойственны два первых процесса.
Отток ионов из листьев. Почти все элементы, за исключением кальция и бора, могут оттекать из листьев, достигших зрелости и начинающих стареть. Среди катионов во флоэмных экссудатах доминирующее место принадлежит калию, на долю которого может приходиться около 60 % суммы катионов. Из всех двухвалентных ионов в заметном количестве перемещается по флоэме только магний; возможно, что для этого он включается в состав комплексных соединений. Железо, марганец, цинк и молибден также перемещаются по флоэме, но их концентрация не превышает средний уровень содержания в других клетках.
Среди анионов в ситовидных элементах флоэмы преобладает фосфат. Часть фосфора находится в виде неорганического фосфата, другая связана в органических соединениях (гексозофосфатах, нуклеотидах, нуклеиновых кислотах). Из других анионов в ситовидных элементах всегда обнаруживается Сl-, концентрация которого во многом зависит от общего уровня хлоридов в почве и в самом растении. Нитраты и сульфаты при определенных условиях (высокая обеспеченность источниками питания, замедление фотовосстановительных реакций) также транспортируются с нисходящим током. Однако более типичным является отток из листа азота и серы в составе органических соединений.
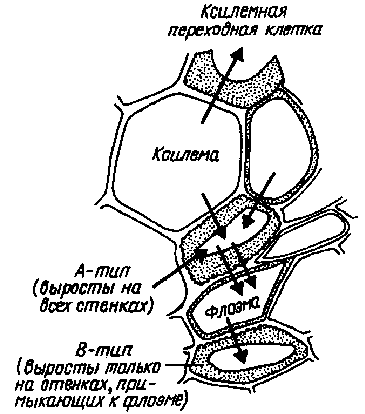
В ситовидные элементы флоэмы ионы могут поступать как путем транспорта их из клеток мезофилла, так и не выходя за пределы листовой жилки, при участии переходных (передаточных) клеток, в стенках которых образуются многочисленные выросты цитоплазмы. Такие лабиринты, значительно увеличивающие контактную поверхность, могут быть поляризованы. Например, у переходных клеток ксилемы (тип А) выросты цитоплазмы есть лишь в стенке, примыкающей к сосуду, что облегчает перенос ионов из ксилемного сока в симпласт ксилемной паренхимы. В переходных клетках, расположенных в жилке между сосудами ксилемы и флоэмы (тип В), выросты имеются на границе с обеими транспортными системами, что обеспечивает эффективный обмен ионами между ними. Нисходящий ток минеральных веществ по флоэме менее значителен по своим масштабам, чем восходящий транспорт по ксилеме. В совокупности они обеспечивают круговорот, способствующий перераспределению элементов в растении.
Дата добавления: 2015-02-03; просмотров: 12483;