Дыхание и обмен веществ в растительной клетке
Каждый механизм накопления энергии вносит определенный, но неравнозначный вклад в общий фонд образования АТФ в клетке. Разные пункты сопряженного фосфорилирования в ЭТЦ также неравноценны с точки зрения энергетической эффективности. При величине Р : О = 3 около 2/3 всего количества АТФ, образуемого при аэробном дыхании, должно поставляться вторым и третьим фосфорилированием дыхательной цепи, т.е. переносом электронов, связанным с цитохромной системой. Из основных субстратов только янтарная кислота и ацетил-КоА окисляются, минуя НАД+ и первую точку фосфорилирования в ЭТЦ.
За счет субстратного фосфорилирования в цикле Кребса образуются только две молекулы АТФ, за счет коферментного - 34. В ходе гликолитического фосфорилирования за вычетом затрат на запуск гликолиза образуются две молекулы АТФ, за счет коферментного - 34. Хотя вклад гликолиза как анаэробной фазы в общий фонд энергии клетки ниже, чем вклад аэробной фазы, гликолиз является одним из важных поставщиков энергии при дефиците кислорода в ткани. Кроме того, факторы, нарушающие фосфорилирование в митохондриях, не затрагивают гликолитической энергетической системы.
Роль гликоза в синтезе АТФ возрастает и тогда, когда энергия ЭТЦ дыхания используется не для фосфорилирования, а на эндэргонические функции митохондрий (активный транспорт ионов, конформационные изменения митохондрий и др.). Значение гликолиза как поставщика энергии заключается еще в том, что биосинтетические процессы, локализованные в цитоплазме, легче используют АТФ, образующуюся в гликолитическом фосфорилировании непосредственно в цитоплазме.
Таким образом, гликолиз, как и субстратное фосфорилирование, не требует высокой степени структурной организации фосфорилирующих механизмов, которые необходимы сопряженному фосфорилированию в ЭТЦ. Поэтому они не нарушаются при повреждении структуры митохондрий. Кроме того, субстратное фосфорилирование нечувствительно к факторам, угнетающим коферментное фосфорилирование, например, при разрушении митохондрий. Вышесказанное позволяет считать гликолиз и субстратное фосфорилирование более надежными источниками энергии в клетке, особенно в неблагоприятных условиях выращивания. Однако для оптимального функционирования клетки важны все механизмы образования макроэргов, поскольку растительные организмы произрастают в изменяющихся условиях среды. Например, существенные изменения в соотношении между отдельными путями дыхания наступают у растений под влиянием фотопериодического воздействия. Так, длинный день активирует аэробные превращения дыхательного субстрата, а короткий день - гликолитические процессы и пентозофосфатный цикл. В период роста листа растяжением скорость гликолиза и цитохромного пути снижается, а скорость альтернативного пути остается постоянной.
Роль дыхания в биосинтетических процессах.Тесная связь дыхания с биосинтетическими функциями клетки не ограничивается использованием реализуемой в ходе окислительных процессов химической энергии дыхательного субстрата. Промежуточные продукты дыхания могут использоваться в процессах новообразования компонентов протоплазмы. Например, аминирование α-кетоглутаровой кислоты приводит к образованию глутаминовой кислоты.

К аминированию путем непосредственного присоединения аммиака способны также ПВК и ЩУК. Так, прямое аминирование ПВК приводит к образованию аланина.
В процессах восстановительного биосинтеза используется почти исключительно НАДФ-Н, тогда как НАД-Н потребляется преимущественно для генерирования АТФ.
С окислительными превращениями трикарбоновых кислот в цикле Кребса тесно связаны процессы биосинтеза жиров. Конденсация двух молекул уксусной кислоты при участии КоА приводит к образованию ацетоуксусной кислоты, при восстановлении которой получается масляная кислота.
Путем последовательного присоединения ацетильных радикалов, активируемых КоА, и происходит наращивание углеродной цепи - синтез жирных кислот в организме.
В ходе гексозомонофосфатного дыхания образуются активные формы пентоз, участвующие в построении важнейших соединений клетки. У гетеротрофных организмов гексозомонофосфатное Дыхание представляет единственный путь в образовании пентоз.
Таким образом, промежуточные продукты дыхания могут дать начало всем основным компонентам растительной клетки, и дыхание в этом случае выполняет функции, которые по своему значению для синтетической функции клетки не уступают фотосинтезу. Далее эритрозо-4-фосфат, образующийся на одном из этапов пентозофосфатного цикла, образует в результате взаимодействия с ФЕП циклическое соединение - шикимовую кислоту. Последняя составляет материальную основу для синтеза всех ароматических аминокислот и в их числе триптофана, который, в свою очередь, является предшественником гетероауксина - основного ростактивирующего гормона растительной ткани.
Шикимовая кислота после ряда превращений может дать начало ароматическим аминокислотам - фенилаланину и тирозину, последующее дезаминирование которых приводит к образованию веществ вторичного происхождения - коричных кислот и кумаринов. С синтезом последних, в свою очередь, связано образование полифенолов. При декарбоксилировании аминокислот образуются амины, являющиеся предшественниками алкалоидов. Важная роль принадлежит дыханию в биосинтезе зеленых пигментов листа, что свидетельствует о тесной связи фотосинтеза и дыхания.
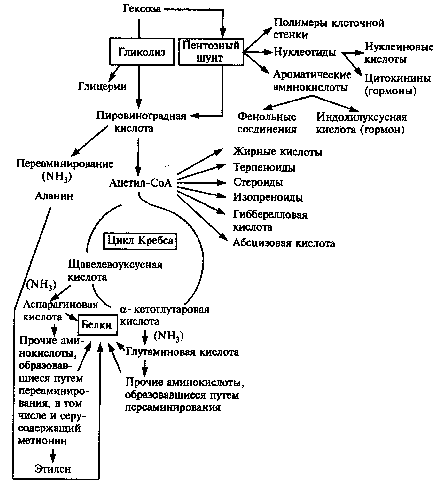
Из изложенного видно, что окислительные превращения дыхательного субстрата занимают важное место в процессах биосинтеза не только белков, жиров и углеводов, но и регуляторов процессов обмена - ферментов и коферментов, веществ вторичного происхождения и гормонов.
Связь дыхания и фотосинтеза. Взаимосвязь дыхания и фотосинтеза - одна из важных и вместе с тем сложных проблем физиологии растений. Хотя при фотосинтезе растение использует конечные продукты дыхания, а субстратами дыхания служат продукты фотосинтеза, эти два процесса в течение длительного периода рассматривали как диаметрально противоположные. Действительно, при фотосинтезе происходит синтез органического вещества, при дыхании - его распад, при фотосинтезе осуществляется восстановление, а при дыхании - окисление вещества. АТФ, образуемый в митохондриях, используется практически на все внутриклеточные процессы, а АТФ хлоропластов - главным образом на процессы ассимиляции СО2. Кроме того, при дыхании укорочение углеродной цепи происходит в результате декарбоксилирования веществ, а при фотосинтезе - карбоксилирования.
Однако, несмотря на все различия, фотосинтез и дыхание тесно связаны. Например, если рассматривать на молекулярном уровне одинаковые начальные и конечные этапы цикла Кальвина и пентозофосфатного пути, многие промежуточные продукты и каталитические системы тождественны. Промежуточные продукты обоих процессов широко используются в различных биосинтезах. Основными функциями как митохондрий, так и хлоропластов являются транспорт электронов и сопряженные с переносом электронов процессы связывания энергии в видеАТФ при фосфорилировании.
При фотосинтезе активирование электрона происходит за счет энергии кванта света, при дыхании - за счет энергии химических связей, сосредоточенной в молекуле дыхательного субстрата. Однако свободная энергия любого органического соединения генетически связана с фотосинтезом. Следовательно, как при фотосинтезе, так и при дыхании источник образования активного электрона - это электромагнитная энергия кванта света.
Много общего обнаруживают компоненты ЭТЦ митохондрий и хлоропластов. Последнее позволяет считать, что при формировании каталитических систем аэробного дыхания, возникших в эволюционном отношении вслед за фотосинтезом, были широко использованы важнейшие компоненты ЭТЦ фотосинтеза.
Ультраструктура и химический состав органелл не только принципиально близки, но и во многих отношениях общие. Обе органеллы способны самостоятельно осуществлять синтез белков. Это свидетельствует о большой автономности этих органелл и способности к самовоспроизведению, подтверждает идею об их происхождении от общего предшественника - пропластиды. Кроме того, практически все продукты процесса гликолиза, цикла Кребса и гексозомонофосфатного пути обычно обнаруживаются в группе соединений, относящихся к самым ранним продуктам фотосинтеза.
Все это свидетельствует о несостоятельности попыток противопоставить дыхание фотосинтезу с общебиологической точки зрения. Как отмечал П.С. Беликов (1980), гликолиз - анаэробное дыхание - филогенетически, вероятно, был первым поставщиком энергии для клетки. Фотосинтез, появившийся в эволюции позднее, обогатил атмосферу кислородом, в результате чего стало возможным аэробное дыхание. Пентозофосфатный цикл, проходящий в условиях высокого содержания кислорода, мог возникнуть еще позднее.
Тесная связь между фотосинтезом и дыханием сохраняется также на уровне целого растения. С тех пор как П. Бойсен-Йенсен (1932) ввел понятие «нетто-ассимиляция», а А.Я. Кокин составил известное уравнение: «Прирост = фотосинтез - дыхание», последнее стало рассматриваться главным образом как растрата ассимилятов. В этот период считалось, что уменьшение дыхания приводит к повышению продуктивности растения.
Концепция «медленное дыхание - высокая продуктивность» обязана своим происхождением обнаруженной среди зерновых культур отрицательной корреляции между интенсивностью дыхания корней и скоростью роста при дефиците влаги. Сорта с низкой ИД корней имели, как правило, более высокую продуктивность при одинаковой ИФ. Однако позже было отмечено, что высокая активность дыхания может компенсироваться увеличением фотосинтеза, а также ускорением роста и развития растений. Подобная неоднозначная оценка роли дыхания подтверждает высказывание В.А. Бриллиант о том, что вопрос о соотношении фотосинтеза и дыхания в процессе создания урожая выходит далеко за рамки простого сопоставления функций синтеза и распада органических соединений растения.
Действительно, скорость как дыхания, так и фотосинтеза может служить показателем функциональной стойкости растений, и их измерение позволяет выявить причины изменений продуктивности в конкретных условиях среды. По мнению О.В. Заленского (1977), в ряде случаев по изменениям фотосинтеза и дыхания можно предсказать не только направление изменений продуктивности растений, но и ее величину.
В конце 60-х годов для посевов многих сельскохозяйственных культур (пшеница, рис, люцерна, хлопчатник и др.) было установлено, что дыхание возрастает не прямо пропорционально увеличению листового индекса или сырой массы растения, как считали ранее.
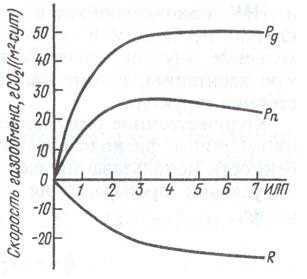
При определенной величине листового индекса кривая дыхания, как и фотосинтеза, выходит (или почти выходит) на плато. Скорость накопления биомассы также не имеет максимума и, следовательно, нет оптимального ИЛП. Эти данные привели исследователей к мысли о тесной связи фотосинтеза и дыхания посева, что количественно было впервые выражено голландским ученым Мак Кри в 1970 г.: R = aPq + bW, где R и Pq — соответственно скорость дыхания и суммарный фотосинтез единицы площади посева, г СО2/(м2 ·сут); W— сухая биомасса, г, выраженная в эквивалентах СО2; а и b - коэффициенты дыхания соответственно на рост и на поддержание.
Таким образом, дыхание посева состоит из дыхания на рост (aPq) и дыхания на поддержание (bW).
Коэффициент дыхания на рост изменяется в небольших пределах (0,2-0,3), гораздо шире изменяется коэффициент дыхания на поддержание - 0,01-0,04, имеющий размерность скорости — г/(г·сут). Величины коэффициентов указывают на то, что в суточном балансе углерода от 20 до 30 % гросс-фотосинтеза тратится на дыхание роста и 1-4 % уже накопленной биомассы - на дыхание поддержания. Дыхание на рост обеспечивает энергией синтез новой биомассы, т.е. превращение продуктов фотосинтеза, например глюкозы, в белки, жиры, липиды и углеводы.
Круг процессов, именуемых термином «поддержание», четко не очерчен. В целом они связаны с энергией, затрачиваемой на ресинтез ферментативных и структурных белков, липидов, РНК и ДНК, обновляющихся в ходе обмена веществ; поддержание в клетках необходимой концентрации ионов и величины рН; сохранение внутриклеточного фонда метаболитов; физиологическую адаптацию, обеспечивающую функционально активное состояние структур.
Количественное разделение дыхания на компоненты и оценка их величин с физиологических позиций позволяют выявить особенности использования ассимилятов растениями в зависимости от условий произрастания. Это хорошо видно на следующей схеме:
Гросс-фотосинтез растения (Pq), суммированный в течение фотопериода, формирует определенное количество субстрата (ΔS), часть которого (ΔSm) используется для генерации энергии и углеродного скелета, необходимого для поддержания статус-кво клетки. Иными словами, ΔSm = bW. Остальная часть субстрата (ΔSq) расходуется на процессы роста массы (ΔW) и связанное с последним дыхание, называемое дыханием роста (ΔSR или aPq). Разделение на дыхание роста и дыхание поддержания только количественное, так как обе компоненты имеют одинаковую биохимическую основу. Принципиальная разница между ними в том, что дыхание роста не зависит от температуры, но сильно зависит от обеспеченности дыхания субстратом, в то время как дыхание поддержания сильно зависит от температуры, но не зависит непосредственно от поступления субстрата.
Дата добавления: 2015-02-03; просмотров: 3701;