Щитки сегментов, 9 — глаза
размельчается и разминается пища, тогда как прочие членики составляют род щупальца. У представителей некоторых отрядов (скорпионы, лжескорпионы) педипальпы превращены в мощные длинные клешни (см. рис. 389, 390,Ж), у других — похожи на ходные ноги (см. рис. 390, Е, Д; рис. 393).
Остальные 4 пары головогрудных конечностей состоят из 6—7 члеников и играют роль ходных ног. Они заканчиваются коготками.
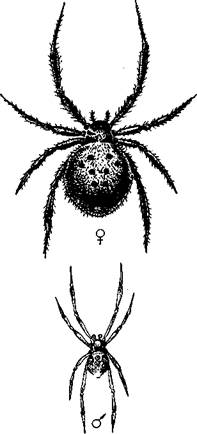
|
| Рис. 391. Самец и самка каракурта Latrodectus tredecimguttatus (по Мариковскому) |
У взрослых паукообразных брюшко лишено типичных конечностей, хотя они, несомненно, произошли от предков, обладавших хорошо развитыми ножками на передних брюшных сегментах. У эмбрионов многих паукообразных (скорпионов, пауков) на брюшке закладываются зачатки ножек, которые лишь впоследствии подвергаются регрессу. Впрочем, и во взрослом состоянии брюшные ножки иногда сохраняются, но в модифицированном виде. Так, у скорпионов на первом сегменте брюшка есть пара половых крышечек, под которыми открывается половое отверстие, на втором — пара гребенчатых органов (см. рис. 389), которые снабжены многочисленными нервными окончаниями и играют роль осязательных придатков. И те и другие представляют видоизмененные конечности. Такова же природа и легочных мешков, расположенных на сегментах брюшка у скорпионов, некоторых пауков и лжескорпионов (с. 413), От конечностей берут свое начало и паутинные бородавки пауков. На нижней поверхности брюшка впереди порошицы у них находится 2—3 пары бугорков, усаженных волосками и несущих труб-ковидные протоки многочисленных паутинных желез (рис. 394). Гомология этих паутинных бородавок брюшным конечностям доказывается не только их эмбриональным развитием, но и строением их у некоторых тропических пауков, у которых бородавки особенно сильно развиты, состоят из нескольких члеников и даже по виду напоминают ножки.
Покровы хелицеровых состоят из кутикулы и подлежащих слоев: гиподермального эпителия (гиподермы) и базальной мембраны. Сама кутикула представляет сложное трехслойное образование. Снаружи располагается липопротеиновый слой, надежно предохраняющий организм от потери влаги при испарении. Это позволило хелицеровым стать настоящей сухопутной группой и заселить самые засушливые районы земного шара. Прочность кутикуле придают белки, задубленные фенолами и инкрустирующие хитин.
|
|
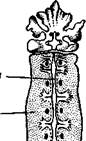
Производными кожного эпителия являются некоторые железистые образования, в том числе ядовитые и паутинные железы. Первые свойственны паукам, жгу-
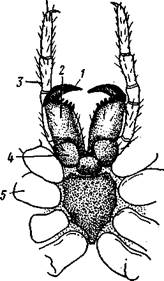
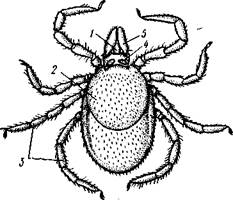
Рис. 392. Собачий клещ Ixodes п-cinus. A — самка, вид со спинной стороны (по Померанцеву); Б — ротовые органы со спинной стороны; В — то же, с брюшной стороны (из Догеля):
/ — хоботок, 2 — туловище, 3 — ходные ноги, 4 — основание хоботка, 5 — педи-пальпа, 6 — гипостом, вырост основания хоботка, 7 — хслицеры. 8 — кроющие пластинки
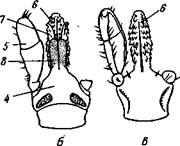
Рис. 393. Ротовые органы паука крестовика Araneus diade-matus (по Иванову):
1 — конечный когтевидный членик хелицеры, 2 — основной членик хе-лицеры, 3 — педнпальпа, 4 — жевательный вырост основного членика педипальпы, 5 — основной членик ходной ноги
|
| 13 |
| 8 |
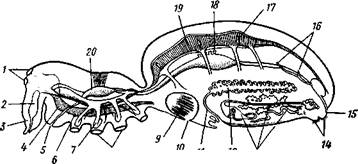
Рис. 394. Схема организации паука (отр. Aranei) (из Аверинцева): I — глаза, 2 — ядовитая железа, 3 — хелицера, 4 — мозг, 5 — рот, 6 — под-глоточный нервный узел, 7 — железистый вырост кишечника, S — основания ходных ног, 9 — легкое, 10 — легочное отверстие — дыхальце, II — яйцевод, 12 — яичник, 13 — паутинные железы, 14 — паутинные бородавки, 15 — анус, 16 — мальпигиевы сосуды. /7 — остии, 18 — протоки печени, 19 — сердце, 10 — глотка, связанная со стенкой тела мускулатурой
топогим и скорпионам; вторые — паукам, лжескорпионам и некоторым клещам.
|
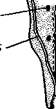
Пищеварительная система у представителей разных отрядов хелице-ровых сильно варьирует. Передняя кишка обыкновенно образует расширение— снабженную сильными мышцами глотку, которая служит в качестве насоса, втягивающего полужидкую пищу, так как паукообразные не принимают твердую пищу кусками. В переднюю кишку открывается пара небольших «слюнных желез». У пауков секрет этих желез и печени способен энергично расщеплять белки. Он вводится в тело убитой добычи и приводит содержимое ее в состояние жидкой кашипы, которая затем всасывается пауком. Здесь имеет место так называемое внекишечное пищеварение.
| II |
|
У большинства паукообразных средняя кишка образует длинные боковые выпячивания, увеличивающие вместимость и всасывающую поверхность кишечника. Так, у пауков (рис. 394) от головогрудной части средней кишки к основаниям конечностей идут 5 пар слепых железистых мешков; аналогичные выпячивания имеются у клещей, сенокосцев и других паукообразных. В брюшной отдел средней кишки открываются протоки парной пищеварительной железы — печени (рис. 395); она выделяет пищеварительные ферменты и служит для всасывания питательных веществ. В клетках печени происходит внутриклеточное пищеварение.
Огромное большинство Arachnida — хищники, но имеются паразитические формы, питающиеся кровью позвоночных, и много растительноядных форм, сосущих соки растений или поедающих растительные остатки.
Выделительная система паукообразных по сравнению с мечехвостами имеет совершенно иной характер. На границе между средней и задней кишкой в пищеварительный канал открывается пара большей частью ветвящихся мальпигиевых сосудов. В отличие от Тга-cheata они энтодермального происхождения, т. е. образуются за счет средней кишки (см. рис. 394, 395). Как в клетках, так и в просвете мальпигиевых сосудов есть многочисленные зерна гуанина — главного продукта выделения паукообразных. Гуанин, как и мочевая кислота, экскретируемая насекомыми (с. 352), обладает малой растворимостью и удаляется из организма в виде кристаллов. Потери влаги при этом минимальные (с. 353), что важно для животных, перешедших к жизни на суше.
| Рис. 395. Пищева рительная система скорпиона (по Павловскому): / — средняя кншка, 2 — печень. Я — протоки печени, 4 — дор-зо-вентральные мышцы, пронизывающие печень, S — мальпн-гиев сосуд, в — задняя кишка |
Кроме мальпигиевых сосудов паукообразные обладают еще и типичными коксальными железами — парными мешковидными образованиями мезодермальной природы, лежащими в двух (реже в одном) сегментах головогруди. Они хорошо развиты у зародышей и в молодом возрасте, но у взрослых животных более или менее атрофируются. Вполне сформированные кок-сальные железы (рис. 396) состоят из концевого эпителиального мешочка, петлевидно извитого канала и более прямого выводного протока с мочевым пузырь-
ком и наружным отверстием. Концевой мешочек отвечает ресничной воронке целомодукта, отверстие которой замкнуто остатком целомическо-го эпителия. Коксальные железы открываются у основания 3-й или 5-й пары конечностей.
|
Нервная система Arachnida разнообразна. Будучи связанной в происхождении с брюшной нервной цепочкой кольчатых червей, у паукообразных она обнаруживает четко выраженную тенденцию к концентрации.
Головной мозг имеет сложное строение. В его состав входят два отдела: передний, иннервирую-щий глаза, — протоцеребрум и задний — тритоцеребрум, посылающий нервы к первой паре конеч-
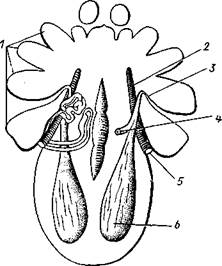
Рис. 396. Коксальные железы сенокосца Garella variegata (из Ланга):
/ — основания ходных ног, 2 — трахейный ствол, 3 — выделительное отверстие, 4 — перерезанный выводной канал железы (на левой стороне он сохранился полностью), 5 — стигма, 6 — резервуар железы (мочевой пузырек)
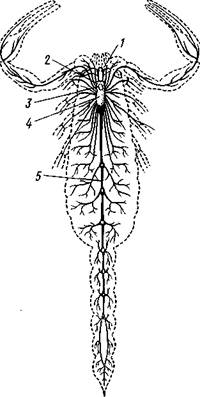
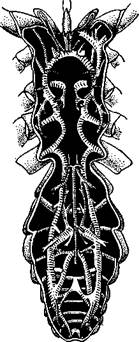
Рис. 397. Нервная система скорпиона Androctonus (из Ганштрома):
/ — глаза, 2 — мозг, 3—окологлоточные
коннективы, 4 — подглоточная ганглиозная
масса, 5 — брюшная нервная цепочка
ностей — хелицерам. Характерный для других членистоногих (ракообразные, насекомые) промежуточный отдел мозга — дейтоцеребрум у паукообразных отсутствует. Связано это с исчезновением у них, как и у остальных хелицеровых, придатков акрона — антеннул, или усиков, которые иннервируются именно от дейтоцеребрума.
Метамерность брюшной нервной цепочки всего яснее сохраняется у скорпионов (рис. 397). У них имеется помимо головного мозга и окологлоточных коннективов большая ганглиозная масса в головогруди на брюшной стороне, дающая нервы к 2—6-й парам конечносте'й и 7 ганглиев, на протяжении брюшного отдела нервной цепочки. У сольпуг кроме сложного головогрудного ганглия на нервной цепочке сохраняется е:ле один узел, а у пауков уже вся цепочка слилась в головогрудный ганглий.
Наконец, у сенокосцев и клещей нет даже явственного разграничения между головным мозгом и головогрудным ганглием (рис. 398), так что нервная система образует вокруг пищевода сплошное ганглиозное кольцо.
|
Органы чувств Arachnida разнообразны. Очень важные для паукообразных механические, осязательные раздражения воспринимаются различно устроенными чувствительными волосками, которые особенно многочисленны на педипальпах. Специальные волоски — трихо-ботрии, расположенные на педипальпах, ногах и поверхности туловища, регистрируют колебания воздуха. Так называемые лировидные органы, представляющие собой небольшие щели в кутикуле, к перепончатому дну которых подходят чувствительные отростки нервных клеток, являются органами химического чувства и служат для обоняния. Органы зрения
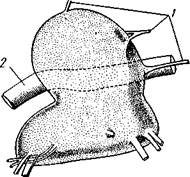
Рис. 398. Центральная нервная система сенокосцев (отр. Opiliones) (по
Гольмгрену): 1 — нервы. 2 — пищевод, пронизывающий
нервную массу
Рис. 399. Трахейная
система сольпуги (из
Ланга)
представлены простыми глазами, которые имеются у большинства паукообразных. Они расположены на дорзальной поверхности головогруди и обычно их бывает несколько: 12, 8, 6, реже 2. У скорпионов, например, имеется пара срединных более крупных глаз и 2—5 пар боковых. У пауков чаще всего 8 глаз, расположенных обычно в две дуги, причем средние глаза передней дуги крупнее остальных.
Скорпионы узнают себе подобных лишь на расстоянии в 2—3 см, а некоторые пауки — за 20—30 см. У пауков-скакунов (сем. Salticidae) зрение играет особенно важную роль: если самцам замазать глаза непрозрачным асфальтовым лаком, то они перестают различать самок и производить характерный для периода спаривания «любовный танец».
Органы дыхания Arachnida разнообразны. У одних — это легочные мешки, у других — трахеи, у третьих—и те и другие одновременно.
Только легочные мешки имеются у скорпионов, жгутоногих и у примитивных пауков. У скорпионов на брюшной поверхности 3—6-го сегментов переднебрюшия располагаются 4 пары узких щелей — дыхалец, которые ведут в легочные мешки (см. рис. 389). В полость мешка вда-
ются многочисленные параллельные друг другу листовидные складки, между которыми остаются узкие щелевидные пространства, в последние проникает через дыхательную щель воздух, а в легочных листочках циркулирует гемолимфа. У жгутоногих и низших пауков имеются лишь две пары легочных мешков.
У большинства других паукообразных (сольпуги, сенокосцы, лжескорпионы, часть клещей) органы дыхания представлены трахеями (рис. 399, 400). На 1—2-м сегментах брюшка (у сольпуг на 1-м членике груди) имеются парные дыхательные отверстия, или стигмы. От каждой стигмы внутрь тела отходит пучок длинных, тонких, слепо замкнутых на концах воздухоносных трубочек эктодермального происхождения (образуются как глубокие впячивания наружного эпителия). У лжескорпионов и клещей эти трубочки, или трахеи, устроены просто и не ветвятся, у сенокосцев они образуют побочные веточки.
Наконец, в отряде пауков оба рода дыхательных органов встречаются совместно. У низших пауков, как уже отмечалось, имеются только легкие; в числе 2 пар они расположены на нижней стороне брюшка. У остальных пауков сохраняется лишь одна передняя пара легких, а позади последних имеется пара трахейных пучков (рис. 400, В), открывающихся наружу двумя стигмами. Наконец, у одного семейства пауков (Caponiidae) легких нет вовсе, а единственные органы дыхания — 2 пары трахей (рис. 400,5).
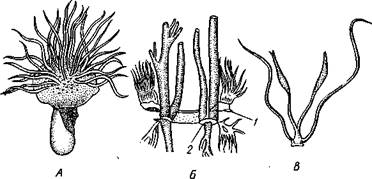
Легкие и трахеи паукообразных возникли независимо друг от друга. Легочные мешки, несомненно, более древние органы. Считается, что раз-

|
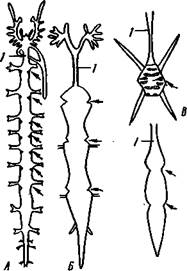
| Рис. 401. Схема происхождения легочных мешков у паукообразных. А — стадия «мечехвоста»; Б — стадия «скорпиона»: / — дыхательная щель легкого. Стрелки показывают токи крови и воздуха |
Рис. 400. Трахеи паукообразных (из Догеля). А — трахеи второй пары Chelifer cancroides (отр. Pseudoscopiones); Б — трахеи паука Nops coccineus — две пары трахейных пучков с соответствующими стигмами (1 и 2); В — трахеи паука Araneus diadematus
витие легких в процессе эволюции было связано с видоизменением брюшных жаберных конечностей, которыми обладали водные предки паукообразных и которые были похожи на жаброносные брюшные ножки мечехвостов. Каждая такая конечность впятилась внутрь тела. При этом образовалась полость для легочных листочков (рис. 401). Боковые края ножки приросли к телу почти на всем протяжении, кроме того
|
| Г |
участка, где сохранилась дыхательная щель. Брюшная стенка легочно-
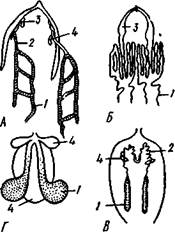
Рис. 402. Строение сердца у паукообразных (из Ланга). А — скорпион; Б — паук; В — клещ; Г —
Дата добавления: 2015-01-24; просмотров: 1490;