Механизмы развития мышечной спастичности
Основным признаком центрального паралича, как и основным фактором, препятствующим восстановлению двигательных функций и значительно ограничивающим жизнедеятельность больного, является спастичность. По определению J. Lance [1980], спастичность - это двигательное нарушение, являющееся одним из компонентов синдрома верхнего мотонейрона и характеризующееся повышением тонических рефлексов на растяжение (или мышечного тонуса) в сочетании с повышением сухожильных рефлексов.
Мышечный тонус определяется двумя факторами:
1) механико-эластическими характеристиками мышечной и соединительной ткани;
2) рефлекторной сократимостью мышц (тоническим рефлексом на растяжение).
Первый фактор, или "внутренняя жесткость" мышечной ткани, имеет несомненное значение в развитии или поддержании спастической гипертонии, поскольку в какой-то степени мышца подобна обыкновенной пружине, возвратная сила действия которой пропорциональная изменению ее длины и в то же время зависит от модуля упругости материала, из которого сделана пружина [Katz R., 1996]. Однако основная роль в поддержании и изменении мышечного тонуса отводится функциональному состоянию сегментарной дуги рефлекса растяжения (миотатического, или проприоцептивного рефлекса). Кратко напомним схему этого рефлекса (рис.4.1. и 4.2).
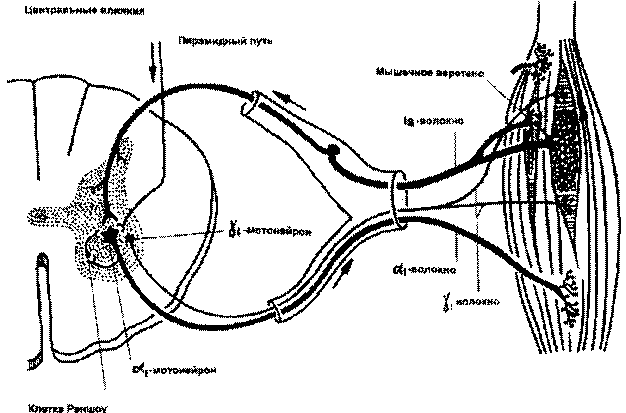
Рис. 4.1. Схема рефлекса поддержания длины мышцы (по П.Дуусу, 1995)
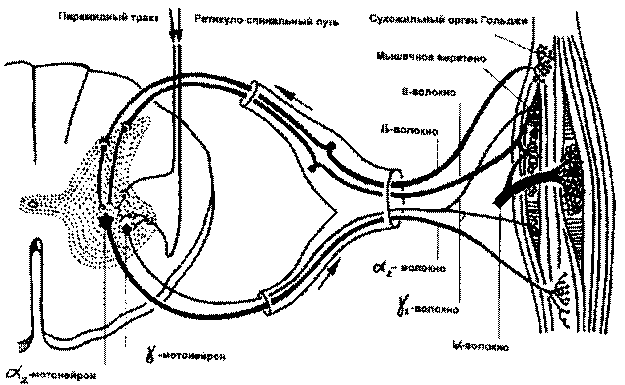
Рис. 4.2. Схема рефлекса поддержания тонуса мышцы (по П.Дуусу, 1995)
Рецепторным элементом миотатического рефлекса является инкапсулированное мышечное веретено (fusus, лат.). Каждая мышца содержит большое количество этих рецепторов. Мышечное веретено состоит из интрафузальных мышечных волокон (в сравнении с обычными экстрафузальными мышечными волокнами они значительно тоньше) и ядерной сумки, оплетенной спиралевидной сетью тонких нервных волокон, представляющих собой первичные чувствительные окончания. На некоторых интрафузальных волокнах имеются еще и вторичные, гроздевидные чувствительные окончания. При растяжении интрафузальных волокон первичные чувствительные окончания усиливают исходящую из них импульсацию, которая через быстро проводящие афферентные волокна типа Iа проводятся к альфа-большим мотонейронам спинного мозга. Оттуда через также быстропроводящие альфа-1 эфферентные волокна импульс идет к экстрафузальным белым мышечным волокнам, которые обеспечивают быстрое (фазическое) сокращение мышцы. От вторичных чувствительных окончаний, реагирующих на тонус мышцы, афферентная импульсация проводится по тонким волокнам II типа через систему вставочных нейронов к альфа-малым мотонейронам, которые иннервируют тонические экстрафузальные мышечные волокна (красные мышечные волокна), обеспечивающие поддержание тонуса и позы.
Интрафузальные волокна иннервируются гамма-нейронами передних рогов спинного мозга. Возбуждение гамма-нейронов, передаваясь по гамма-волокнам к мышечному веретену, сопровождается сокращением полярных отделов интрафузальных волокон и растяжением их экваториальной части, при этом изменяется исходная чувствительность рецепторов к растяжению (происходит снижение порога возбудимости рецепторов растяжения и усиливается тоническое напряжение мышцы).
Гамма-мотонейроны находятся под влиянием центральных (супрасегментарных) воздействий, передающихся по волокнам, которые идут от мотонейронов оральных отделов головного мозга в составе пирамидного, ретикулоспинального, вестибулоспинального трактов. Таким образом, мышечный тонус может регулироваться непосредственно головным мозгом, что очень важно для выполнения произвольных движений. При этом, если роль пирамидной системы заключается в преимущественной регуляции фазических (т.е. быстрых, целенаправленных) компонентов произвольных движений, то экстрапирамидная система обеспечивает плавность произвольных движений, их "настройку" в соответствии с решаемой задачей, т.е. преимущественно регулирует тоническую иннервацию мышечного аппарата.
В регуляции мышечного тонуса принимают участие и тормозные механизмы, которые реализуются с помощью рецепторов Гольджи, расположенных в сухожилиях мыщц, и вставочных клеток Реншоу, находящихся в передних рогах спинного мозга. Сухожильные рецепторы Гольджи при растяжении или значительном напряжении мышцы посылают афферентные импульсы, которые проводятся по быстропроводящим волокнам 1b типа в спинной мозг и оказывают тормозящее воздействие на мотонейроны передних рогов. Вставочные нейроны Реншоу активизируются через коллатерали при возбуждении альфа-мотонейронов и действуют по принципу отрицательной обратной связи, тормозя активность последних. Таким образом, нейрогенные механизмы регуляции мышечного тонуса многообразны и сложны.
Повышение мышечного тонуса при спастических параличах происходит в результате усиления тонического рефлекса на растяжение. В настоящее время спастичность связывают с нарушением не какого-либо одного, а целого ряда нейрофизиологических механизмов, среди которых решающая роль отводится нарушению дифференцированной регуляции альфа- и гамма-мотонейронов, гипервозбудимости спинальных альфа-мотонейронов, уменьшению активности некоторых ингибиторных механизмов [Гехт А.Б. и соавт., 1998]. Спастичные параличи нередко называют "пирамидными", однако в настоящее время полагают, что повышение мышечного тонуса обусловлено поражением не собственно пирамидных волокон, а тесно переплетенных с ними волокон экстрапирамидной системы, в частности, кортико-ретикулярного и вестибулоспинального трактов. При этом среди волокон, контролирующих активность системы "гамма-нейрон - мышечное веретено", в большей степени обычно страдают ингибирующие волокна, тогда как активирующие сохраняют свое влияние на мышечные веретена. Следствием этого является спастичность мышц, гиперрефлексия, появление патологических рефлексов, первоочередная утрата наиболее тонких произвольных движений. Отсутствие или слабая выраженность мышечных атрофии при центральном параличе объясняется тем, что верхний мотонейрон в гораздо меньшей степени влияет на трофику мышц в сравнении с нижним двигательным нейроном.
Степень спастичности чаще всего определяют по модифицированной 5-ти балльной шкале Ашфорта. Объективизацию состояния мышечного тонуса и контроль за динамикой спастической гипертонии возможно проводить с помощью методов биомеханики и электромиографии. Так, проводились попытки оценивать выраженность спастичности по таким электромиографическим показателям, как отношение величины максимального Н-рефлекса к М-ответу (Нmax/Мmax), кривая восстановления Н-рефлекса; однако корреляция этих показателей с клинической выраженностью спастической гипертонии оказалась низкой [Katz R., 1996]. Более информативны биомеханические методы, например, "маятниковый" тест. Этот тест предполагает выведение сустава из состояния равновесия путем дозированного внешнего воздействия с последующей регистрацией возникших колебаний соответствующего сегмента конечности; скорость затухания этих колебаний отражает тонус мышц конечности [Katz R. и соавт., 1992].
Дата добавления: 2015-01-09; просмотров: 1281;