НЕЙРОТРАНСМИТТЕРНАЯ РЕГУЛЯЦИЯ ГИПОФИЗОТРОПНЫХ НЕЙРОСЕКРЕТОРНЫХ КЛЕТОК
Уже упоминалось, что в центральной нервной системе не существует анатомически и гистологически отграниченных областей, которые могли бы считаться регулирующими выделение данного гипофизотропного гормона. Скорее в отношении каждого гормона существует нейротрансмиттерное «кодирование» нейросекреторных клеток, определяющее выделение их специфических продуктов: гипофизотропных гормонов. Это означает, что к выделению определенного гипофизотропного гормона могут приводить посылки из разных клеток относительно широкой анатомической области, если только они реализуются с помощью одного вида нейротрансмиттеров. Кроме того, поскольку нервная система опосредует секрецию гипофизарных гормонов в разных условиях (периодическая, базальная и стрессорная секреция), то вполне может быть, что в каждом из этих случаев регуляция секреции данного гипофизарного гормона осуществляется с помощью разных нейротрансмиттеров. В связи с этим нейросекреторная клетка должна была бы обладать рецепторами к разным нейротрансмиттерам, каждый из которых высвобождается определенными волокнами, активирующимися под действием определенных стимулов. В отношении гипоталамо-гипофизарно-надпочечниковой системы показано, что данный нейротрансмиттер может активировать секрецию АКТГ лишь при имплантации в определенную область гипоталамуса или лимбической системы и что данная гипоталамическая область может стимулироваться (в плане секреции АКТГ) лишь определенным нейротрансмиттером. Нейротрансмиттеры могут оказывать на функцию нейросекреторной клетки либо возбуждающее, либо тормозное влияние. Окончательная реакция данной нейросекреторной клетки, проявляющаяся началом или прекращением секреции, отражает, таким образом, эффекты местной концентрации возбуждающих и тормозных нейротрансмиттеров, модифицированные местной концентрацией различных ионов, рН, а также гормонов периферической железы и гипофиза.
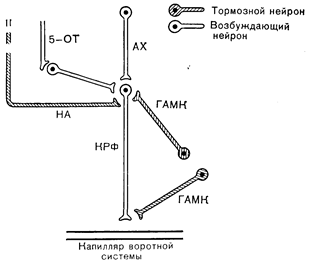
Рис. 6—5. Предполагаемая модель регуляции выделения кортикотропин-рилизинг фактора (КРФ) из гипоталамуса. На модели можно видеть: КРФ-нейрон с окончанием на капилляре воротной системы; 5-ОТ- (окситриптамин) -волокно, стимулирующее выделение КРФ за счет активации холинергического интернейрона (AX — ацетилхолин); конечный общий холинергический путь выделения КРФ; показаны два холинергических нейрона, одни из которых — вставочный нейрон, расположенный между 5-ОТ-волокном и КРФ-клеткой (регуляция циркадного ритма?), а второй не имеет 5-ОТ-синапса и может контролировать стрессорное выделение КРФ; норадренергическое волокно (НА), которое тормозит секрецию КРФ; ГАМК-тормозные нейроны, влияние которых могло бы осуществляться пре- либо постсинаптически (Jones M. Т., Hillhouse E. W., Burden J., Neurotransmitter regulation of corticotropin-releasing factor in vitro Ann. NJ Acad. Press, 1977, 297, 536).
Пытаясь представить себе нейротрансмиттерный контроль выделения гипофизотропных гормонов (рис. 6—5), следует учитывать, что: 1 — такие трансмиттеры могут действовать либо постсинаптически, либо за счет аксоаксональных контактов; 2 — нейротрансмиттерная регуляция выделения гипофизотропных гормонов может осуществляться через мультисинаптические пути (см. раздел «Внегипоталамические афферентные системы»), так что данный нейротрансмиттер может действовать в синапсе, расположенном проксимальнее того, который функционирует на последнем этапе регуляции нейросекреторной клетки; 3 — как уже отмечалось, эффект данного трансмиттера может зависеть от концентрации других нейротрансмиттеров, своей собственной концентрации (разный эффект низкой и высокой концентрации), временной точки циркадного ритма и гормонального и физиологического состояния особи. Определение локализации нейротрансмиттеров в отдельных областях ЦНС трудоемкая задача, требующая методов, которые разработаны пока еще не для всех предполагаемых нейротрансмиттеров. Для того чтобы понять роль данного нейротрансмиттера в регуляции гормональной функции, необходимо также сопоставлять изменения в содержании этого нейротрансмиттера с изменениями в его кругообороте и секреции гормона. В гипоталамусе, и конкретно в срединном возвышении, найдены практически все предполагаемые нейротрансмиттеры: катехоламины, индоламины, ацетилхолин, гистамин и g-аминомасляная кислота (ГАМК), причем здесь же обнаружены и ферменты, принимающие участие в их синтезе и метаболизме.
С помощью гистохимических и химических методик удалось весьма подробно выяснить распределение сетей моноаминергических нейронов в ЦНС крысы. Сходное распределение имеется, по-видимому, и у человека. Прямых способов определения ацетилхолина не существует, представления о его распределении основаны на результатах применения гистологических методик и биохимического определения холинацетилтрансферазы и ацетилхолинэстеразы. Имеются также некоторые авторадиографические доказательства присутствия сетей ГАМК-содержащих нейронов. Их локализация в ЦНС изучена недостаточно.
До настоящего времени нет полного единогласия в отношении специфических эффектов данного нейротрансмиттера на выделение конкретного гипофизотропного гормона. Существующие данные суммированы в табл. 6—2, в которой указаны также противоречия, часть из которых можно было бы объяснить видовыми различиями.
Таблица 6—2. Нейротрансмиттерная регуляция секреции гормонов гипофиза
| ГР | Пролактин | Гонадотропин | ттг | АКТГ | |
| Норадреналин | | — | ? | +¯ | |
| Дофамин | | ¯ | ¯ или | ¯ или | ?¯ |
| Серотонин | | | ?¯ | ? | +¯ |
| ГАМК | | ? | ?¯ | ||
| Гистамин | ? | ||||
| Ацетилхолин | ? | ИЛИ ¯ | |
Примечание. Стрелки, направленные вверх, означают повышение, вниз — снижение секреции: знак минус — отсутствие эффекта, подчеркнутая стрелка —» преимущественный эффект.
Все изложенное исходит из того, что влияние нейротрансмиттеров на секрецию гипофизарных гормонов опосредуется их действием на выделение гипофизотропных гормонов. Однако значительные количества дофамина и серотонина присутствуют в переднем гипофизе. Прямое действие дофамина на секрецию гипофизарных гормонов показано, в частности, в отношении пролактина; не исключено, что другие биогенные амины могут непосредственно влиять на секрецию и других гормонов гипофиза.
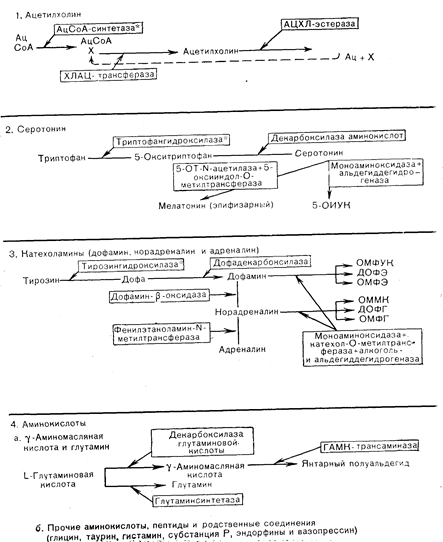
Рис. 6—6. Нейротрансмиттеры в головном мозге млекопитающих; ферменты и главные метаболиты их обмена.
Звездочкой обозначены ферменты, лимитирующие скорость реакции: Ац — ацетат, ацетил или уксусная кислота; ХЛ — холин; 5-ОИУК — 5-оксииндолуксусная кислота- ОМФУК — 4-окси-З-метоксифенилуксусная кислота; ДОФЭ — 3,4-диоксифенилэтанол; ОМФЭ — 4-окси-З-метоксифенилэтанол: ОММК — 4-окси-3-метокси-D-миндальная кислота; ДОФГ — 3,4-диоксифенилгликоль; ОМФГ — 4-окси-3-метоксифенилгликоль.
Дата добавления: 2015-01-19; просмотров: 1734;