Проводящие ткани
Проводящие ткани служат для передвижения по растению растворенных в воде питательных веществ. Они возникли как следствие приспособления растений к жизни на суше. В связи с жизнью в двух средах – почвенной и воздушной, возникли две проводящие ткани, по которым вещества передвигаются в двух направлениях.
По ксилеме от корней к листьям поднимаются вещества почвенного питания – вода и растворенные в ней минеральные соли (восходящий, или транспирационный ток).
По флоэме от листьев к корням передвигаются вещества, образовавшиеся в процессе фотосинтеза, главным образом сахароза (нисходящий ток). Так как эти вещества представляют собой продукты ассимиляции углекислого газа, транспорт веществ по флоэме называют током ассимилятов.
Проводящие ткани образуют в теле растения непрерывную разветвленную систему, соединяющую все органы – от тончайших корешков до самых молодых побегов. Ксилема и флоэма представляют собой сложные ткани, в их состав входят разнородные элементы – проводящие, механические, запасающие, выделительные. Самыми важными являются проводящие элементы, именно они выполняют функцию проведения веществ.
Ксилема и флоэма формируются из одной и той же меристемы и, поэтому, в растении всегда располагаются рядом.Первичные проводящие ткани образуются из первичной латеральной меристемы – прокамбия, вторичные – из вторичной латеральной меристемы – камбия. Вторичные проводящие ткани имеют более сложное строение, чем первичные.
Ксилема (древесина) состоит из проводящих элементов – трахеид и сосудов (трахей), механических элементов -древесинных волокон (волокон либриформа) и элементов основной ткани - древесинной паренхимы.
Проводящие элементы ксилемы носят название трахеальных элементов. Различают два типа трахеальных элементов –трахеиды и членики сосудов (рис. 3.26 ).
Трахеида представляет собой сильно вытянутую в длину клетку с ненарушенными первичными стенками. Передвижение растворов происходит путем фильтрации через окаймленные поры. Сосуд состоит из многих клеток, называемых членикамисосуда. Членики расположены друг над другом, образуя трубочку. Между соседними члениками одного и того же сосуда имеются сквозные отверстия – перфорации. По сосудам растворы передвигаются значительно легче, чем по трахеидам.

Рис. 3.26. Схема строения и сочетания трахеид (1) и члеников сосуда (2).
Трахеальные элементы в зрелом, функционирующем состоянии – мертвые клетки, не имеющие протопластов. Сохранение протопластов затрудняло бы передвижение растворов.
Сосуды и трахеиды передают растворы не только в вертикальном, но и в горизонтальном направлении в соседние трахеальные элементы и в живые клетки. Боковые стенки трахеид и сосудов сохраняются тонкими на большей или меньшей площади. В то же время они имеют вторичные утолщения, придающие стенкам прочность. В зависимости от характера утолщений боковых стенок трахеальные элементы называются кольчатыми, спиральными, сетчатыми, лестничными иточечно-поровыми ( рис. 3.27).
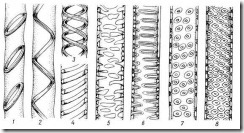
Рис. 3.27. Типы утолщения и поровости боковых стенок у трахеальных элементов : 1 – кольчатое, 2-4 – спиральные, 5 – сетчатое утолщения; 6 – лестничная, 7 – супротивная, 8 – очередная поровость.
Вторичные кольчатые и спиральные утолщения прикрепляются к тонкой первичной стенке посредством узкого выступа. При сближении утолщений и образовании между ними перемычек возникает сетчатое утолщение, переходящее в окаймленные поры. Эту серию (рис. 3.27 ) можно рассматривать как морфогенетический, эволюционный ряд.
Вторичные утолщения клеточных стенок трахеальных элементов одревесневают (пропитываются лигнином), что придает им дополнительную прочность, но ограничивает возможности роста в длину. Поэтому в онтогенезе органа сначала появляются еще способные растягиваться кольчатые и спиральные элементы, не препятствующие росту органа в длину. Когда рост органа прекращается, возникают элементы, неспособные к продольному растяжению.
В процессе эволюции первыми появились трахеиды. Они найдены у первых примитивных наземных растений. Сосуды появились значительно позже путем преобразования трахеид. Сосудами обладают почти все покрытосеменные растения. Споровые и голосеменные растения, как правило, лишены сосудов и обладают только трахеидами. Лишь в виде редкого исключения сосуды встречены у таких споровых, как селагинелла, некоторых хвощей и папоротников, а также у немногих голосеменных (гнетовые). Однако у этих растений сосуды возникли независимо от сосудов покрытосеменных. Возникновение сосудов у покрытосеменных растений означало важное эволюционное достижение, так как облегчило проведение воды; покрытосеменные растения оказались более приспособленными к жизни на суше.
Древесинная паренхима и древесинные волокна выполняют запасающие и опорные функции соответственно.
Флоэма (луб) состоит из проводящих - ситовидных - элементов, сопровождающих клеток (клеток-спутниц), механических элементов – флоэмных (лубяных) волокон и элементов основной ткани – флоэмной (лубяной) паренхимы.
В отличие от трахеальных элементов проводящие элементы флоэмы и в зрелом состоянии остаются живыми, а их клеточные стенки – первичными, неодревесневшими. На стенках ситовидных элементов имеются группы мелких сквозных отверстий –ситовидные поля, через которые сообщаются протопласты соседних клеток и происходит транспорт веществ. Различают два типа ситовидных элементов – ситовидные клетки и членики ситовидных трубок.
Ситовидные клетки являются более примитивными, они присущи споровым и голосеменным растениям. Ситовидная клетка – это одна клетка, сильно вытянутая в длину, с заостренными концами. Ее ситовидные поля рассеяны по боковым стенкам. Кроме того, ситовидные клетки имеют и другие примитивные признаки: они лишены специализированных сопровождающих клеток и в зрелом состоянии содержат ядра.
У покрытосеменных растений транспорт ассимилятов осуществляют ситовидные трубки (рис. 3.28 ). Они состоят из многих отдельных клеток – члеников, расположенных один над другим. Ситовидные поля двух соседних члеников образуютситовидную пластинку. Ситовидные пластинки имеют более совершенное строение, чем ситовидные поля (перфорации крупнее и их больше).
В члениках ситовидных трубок в зрелом состоянии отсутствуют ядра, однако они остаются живыми и деятельно проводят вещества. Важная роль в проведении ассимилятов по ситовидным трубкам принадлежит сопровождающим клеткам (клеткам-спутницам). Каждый членик ситовидной трубки и его сопровождающая клетка (или две-три клетки в случае дополнительного деления) возникают одновременно из одной меристематической клетки. Клетки–спутницы имеют ядра и цитоплазму с многочисленными митохондриями; в них происходит интенсивный обмен веществ. Между ситовидными трубками и прилегающими к ним сопровождающими клетками имеются многочисленные цитоплазматические связи. Считается, что клетки-спутницы вместе с члениками ситовидных трубок составляют единую физиологическую систему, осуществляющую ток ассимилятов.
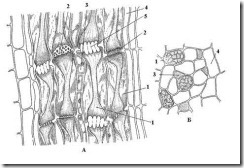
Рис. 3.28. Флоэма стебля тыквы на продольном (А) и поперечном (Б) срезе : 1 – членик ситовидной трубки; 2 – ситовидная пластинка; 3 – сопровождающая клетка; 4 – лубяная (флоэмная) паренхима; 5 – закупоренная ситовидная пластинка.
Длительность функционирования ситовидных трубок невелика. У однолетников и в надземных побегах многолетних трав – не более одного вегетационного периода, у кустарников и деревьев – не более трех-четырех лет. При отмирании живого содержимого ситовидной трубки, отмирает и клетка-спутница.
Лубяная паренхима состоит из живых тонкостенных клеток. В ее клетках часто накапливаются запасные вещества, а также смолы, танниды и др. Лубяные волокна играют опорную роль. Они присутствуют не у всех растений.
В теле растения ксилема и флоэма расположены рядом, образуя или слои, или обособленные тяжи, которые называютпроводящими пучками. Различают несколько типов проводящих пучков (рис. 3.29 ).
Закрытые пучки состоят только из первичных проводящих тканей, они не имеют камбия и далее не утолщаются. Закрытые пучки характерны для споровых и однодольных растений. Открытые пучки имеют камбий и способны к вторичному утолщению. Они характерны для голосеменных и двудольных растений.
В зависимости от взаимного расположения флоэмы и ксилемы в пучке различают следующие типы. Наиболее обычныколлатеральные пучки, в которых флоэма лежит по одну сторону от ксилемы. Коллатеральные пучки могут быть открытыми (стебли двудольных и голосеменных растений) и закрытыми (стебли однодольных растений). Если с внутренней стороны от ксилемы располагается дополнительно тяж флоэмы, такой пучок называется биколлатеральным. Биколлатеральные пучки могут быть только открытыми, они характерны для некоторых семейств двудольных растений (тыквенные, пасленовые и др.).
Встречаются также концентрические пучки, в которых одна проводящая ткань окружает другую. Они могут быть только закрытыми. Если в центре пучка находится флоэма, а ксилема ее окружает, пучок называется центрофлоэмным, илиамфивазальным. Такие пучки часто встречаются в стеблях и корневищах однодольных растений. Если в центре пучка располагается ксилема, и ее окружает флоэма, пучок называется центроксилемным, или амфикрибральным. Центроксилемные пучки обычны у папоротников.
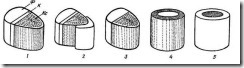
Рис. 3.29. Типы проводящих пучков : 1 – открытый коллатеральный; 2 – открытый биколлатеральный; 3 – закрытый коллатеральный; 4 – концентрический закрытый центрофлоэмный; 5 – концентрический закрытый центроксилемный; К – камбий; Кс – ксилема; Ф – флоэма.
Многие авторы выделяют радиальные пучки. Ксилема в таком пучке располагается в виде лучей от центра по радиусам, а флоэма – между лучами ксилемы. Радиальный пучок – характерный признак корня первичного строения.
5.Механи́ческая, запасающая, воздухоносная ткани. Строение, функции
Механи́ческая ткань — вид ткани в растительном организме, волокна из живых и мёртвых клеток с сильно утолщённой клеточной стенкой, придающие механическую прочность организму. Возникает из верхушечной меристемы, а также в результате деятельности прокамбия и камбия.
Степень развития механических тканей во многом зависит от условий, они почти отсутствуют у растений влажных лесов, у многих прибрежных растений, но зато хорошо развиты у большинства растений засушливых местообитаний.
Механические ткани присутствуют во всех органах растения, но наиболее они развиты по перифериистебля и в центральной части корня.
Выделяют следующие типы механических тканей:
колленхима — эластичная опорная ткань первичной коры молодых стеблей двудольных растений, а также листьев. Состоит из живых клеток с неравномерно утолщёнными неодревесневшими первичными оболочками, вытянутыми вдоль оси органа. Создаёт опору растению.
склеренхима — прочная ткань из быстро отмирающих клеток с одревесневшими и равномерно утолщёнными оболочками. Обеспечивает прочность органов и всего тела растений. Различают два типа склеренхимных клеток:
волокна — длинные тонкие клетки, обычно собранные в тяжи или пучки (например, лубяные илидревесинные волокна).
склереиды — округлые мёртвые клетки с очень толстыми одревесневшими оболочками. Ими образованы семенная кожура, скорлупа орехов, косточки вишни,сливы, абрикоса; они придают мякоти груш характерный крупчатый характер. Встречаются группами в корке хвойных и некоторых лиственных пород, в твердых оболочках семян и плодов. Их клетки круглой формы с толстыми стенками и маленьким ядром.
Механические ткани обеспечивают прочность органов растений. Они составляют каркас, поддерживающий все органы растений, противодействуя их излому, сжатию, разрыву. Основными характеристиками строения механических тканей, обеспечивающими их прочность и упругость, являются мощное утолщение и одревеснение их оболочек, тесное смыкание между клетками, отсутствие перфораций в клеточных стенках.
Механические ткани наиболее развиты в стебле, где они представлены лубяными и древесинными волокнами. В корнях механическая ткань сосредоточена в центре органа.
В зависимости от формы клеток, их строения, физиологического состояния и способа утолщения клеточных оболочек различают два вида механической ткани: колленхиму и склеренхиму, (рис. 8.4).

Рис. 8.4. Механические ткани: а —уголковая колленхима; 6— склеренхима; в -— склереиды из плодов алычи: 1 —цитоплазма, 2 —утолщенная клеточная стенка, 3 — поровые канальцы.
Колленхима представлена живыми паренхимными клетками с неравномерно утолщенными оболочками, делающими их особенно хорошо приспособленными для укрепления молодых растущих органов. Будучи первичными, клетки колленхимы легко растягиваются и практически не мешают удлинению той части растения, в которой находятся. Обычно колленхима располагается отдельными тяжами или непрерывным цилиндром под эпидермой молодого стебля и черешков листьев, а также окаймляет жилки в листьях двудольных. Иногда колленхима содержит хлоропласты.
Склеренхима состоит из вытянутых клеток с равномерно утолщенными, часто одревесневшими оболочками, содержимое которых отмирает на ранних стадиях. Оболочки склеренхимных клеток обладают высокой прочностью, близкой к прочности стали. Эта ткань широко представлена в вегетативных органах наземных растений и составляет их осевую опору.
Различают два типа склеренхимных клеток: волокна и склереиды. Волокна — это длинные тонкие клетки, обычно собранные в тяжи или пучки (например, лубяные или древесинные волокна). Склереиды — это округлые мертвые клетки с очень толстыми одревесневшими оболочками. Ими образованы семенная кожура, скорлупа орехов, косточки вишни, сливы, абрикоса; они придают мякоти груш характерный крупчатый характер.
Основная ткань, или паренхима, состоит из живых, обычно тонкостенных клеток, которые составляют основу органов (откуда и название ткани). В ней размещены механические, проводящие и другие постоянные ткани. Основная ткань выполняет ряд функций, в связи с чем различают ассимиляционную (хлоренхиму), запасающую, воздухоносную (аэренхиму) и водоносную паренхиму (рис. 8.5).

Рис 8.5. Паренхимные ткани: 1—3 — хлорофиллоносная (столбчатая, губчатая и складчатая соответственно); 4—запасающая (клетки с зернами крахмала); 5 — воздухоносная, или аэренхима.
В клетках запасающей паренхимы откладываются белки, углеводы и другие вещества. Она хорошо развита в стеблях древесных растений, в корнеплодах, клубнях, луковицах, плодах и семенах. У растений пустынных местообитаний (кактусы) и солончаков в стеблях и листьях имеется водоносная паренхима, служащая для накопления воды (например, у крупных экземпляров кактусов из рода карнегия в тканях содержится до 2—3 тыс. л воды). У водных и болотных растений развивается особый тип основной ткани — воздухоносная паренхима, или аэренхима. Клетки аэренхимы образуют крупные воздухоносные межклетники, по которым воздух доставляется к тем частям растения, связь которых с атмосферой затруднена.

|
| Аэренхима в стебле Schoenoplectus tabernaemontani |
Аэренхима (или Эренхима) — воздухоносная ткань у растений, построенная из клеток, соединённых между собой так, что между ними остаются крупные заполненные воздухом пустоты (крупные межклетники).
В некоторых руководствах аэренхиму рассматривают как разновидность основной паренхимы.
Построена аэренхима бывает либо из обыкновенных паренхимных клеточек, либо из клеток звёздчатой формы, соединённых друг с другом своими отрогами. Характеризуется наличием межклетников.
Назначение.Встречается такая воздухоносная ткань у водных и болотных растений, и назначение её двоякое. Прежде всего — это вместилище запасов воздуха для потребностей газового обмена. У растений, целиком погружённых в воду, условия газового обмена гораздо менее удобны, чем у наземных растений. Тогда как последние окружены со всех сторон воздухом, водные растения в лучшем случае находят в окружающей среде очень небольшие его запасы; эти запасы поглощаются уже поверхностными клетками, а в глубину толстых органов уже не доходят. Обеспечить себе нормальный газовый обмен растение может при этих условиях двумя путями: либо увеличивая поверхность своих органов при соответствующем уменьшении их массивности, либо собирая внутри своих тканей запасы воздуха. Оба эти способа и наблюдаются в действительности.
Газообмен.С одной стороны, у многих растений подводные листья чрезвычайно сильно рассечены, как, например, у водяного лютика (англ.)русск. (Ranunculus aquatilis), Ouvirandrafene s tralis и проч.
С другой стороны, в случае массивности органов, они представляют собой рыхлую, наполненную воздухом губчатую массу. В течение дня, когда, благодаря процессу ассимиляции, растение выделяет кислорода во много раз больше, чем это необходимо для целей дыхания, выделенный кислород и собирается про запас в крупных межклетниках аэренхимы. В солнечную погоду значительные количества выделенного кислорода не помещаются в межклетниках и выходят наружу сквозь различные случайные отверстия в тканях. С наступлением ночи, когда процесс ассимиляции прекращается, запасённый кислород потребляется постепенно на дыхание клеток, а взамен его в воздухоносные полости аэренхимы выделяется клетками углекислота, чтобы в свою очередь днём пойти на нужды ассимиляции. Так днём и ночью продукты жизнедеятельности растения, благодаря присутствию аэренхимы, не растрачиваются понапрасну, а оставляются про запас, чтобы быть использованными в следующий период деятельности.
Что касается растений болотных, то в особо невыгодных условиях в смысле дыхания находятся у них корни. Под слоем воды, в пропитанной водою почве происходят разного рода процессы брожения и гниения; кислород в самых верхних слоях почвы уже нацело поглощён, дальше создаются уже условия анаэробной жизни, протекающей в отсутствие кислорода. Корни болотных растений не могли бы существовать при таких условиях, не будь у них в распоряжении запаса воздуха в аэренхиме.
Отличие болотных растений и не вполне погружённых водных растений от вполне погружённых состоит в том, что обновление газов внутри аэренхимы происходит не только благодаря жизнедеятельности тканей, а и при помощи диффузии (и термодиффузии); в наземных органах система межклетников открывается наружу массой мельчайших отверстий — устьиц, сквозь которые путем диффузии и уравнивается по составу воздух межклетников с окружающим воздухом. Однако при очень крупных размерах растения такой путь обновления воздуха в аэренхиме корней был бы недостаточно быстр. Соответственно этому, например, у мангровых деревьев, растущих по морским берегам с илистым дном, некоторые разветвления корней растут из ила вверх и выносят в воздух, над поверхностью воды, свои верхушки, поверхность которых пронизана многочисленными отверстиями. Такие «дыхательные корни» имеют целью более быстрое обновление воздуха в аэренхиме питающих корней, разветвленных в бескислородном иле морского дна.
Уменьшение удельного веса
Второй задачей аэренхимы является уменьшение удельного веса растения. Тело растения тяжелее воды; аэренхима играет для растения роль плавательного пузыря; благодаря её присутствию даже тонкие, бедные механическими элементами органы держатся прямо в воде, а не падают в беспорядке на дно. Поддержание органов, главным образом листьев, в положении, благоприятном для жизненных отправлений растения, достигаемое у наземных растений дорогой ценой образования массы механических элементов, достигается здесь у водяных растений просто переполнением аэренхимы воздухом.
Особенно ясно выражена эта вторая задача аэренхимы у плавающих листьев, где запросы дыхания могли бы быть удовлетворены и без помощи аэренхимы. Благодаря обилию межклетных воздухоносных ходов, лист не только плавает на поверхности воды, но и способен выдержать некоторую тяжесть. Особенно славятся этим свойством гигантские листья Victoria regia. Аэренхима, выполняющая роль плавательных пузырей, нередко и образует в самом деле на растении пузыревидные вздутия. Такие пузыри встречаются как у цветковых растений (Eichhornia crassipes, Trianea bogotensis), так и у высших водорослей: Sargassum bacciferum. Fucus vesiculosus и другие виды снабжены прекрасно развитыми плавательными пузырями.
Дата добавления: 2015-01-19; просмотров: 5052;