Дренажный аппарат
Дренажный аппарат состоит из:
1) внутренней склеральной борозды;
2) трабекулярной сети;
3) шлеммова и коллекторных каналов.
Внутренняя склеральная борозда (склеральный валик)представляет собой расположенное циркулярно углубление на внутренней поверхности лимба (рис. 3.3.4—3.3.6). Задней границей внутренней склеральной борозды являются пучки циркулярным образом расположенных коллагеновых волокон, которые формируют склеральную шпору или задний пограничный круг Швальбе. В борозде снаружи размещается шлеммов канал, а кнутри — «корнеосклеральная часть» трабекулярной сети (рис. 3.3.4—3.3.6).

Рис. 3.3.5. Изменение проходимости дренажной системы при расслаблении (а) и сокращении (б) ресничной мышцы:
сокращение мышцы приводит к ее утолщению, что сопровождается уменьшением пространства между мышечными волокнами и уменьшением объема увеасклерального пути оттока. В то же время сокращение мышцы приводит к натяжению склеральной шпоры и расширению, пространств между трабекулами, что способствует уменьшению резистентности трабекулярной сети оттоку камерной влаги

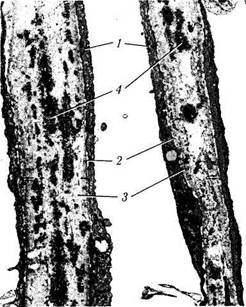
Рис. 3.3.6. Строение дренажной системы при использовании сканирующей электронной микроскопии (по Fine, Yanoff, 1972):
I — роговица; 2 — задняя поверхность роговицы; 3 — коллекторный канал; 4 — шлеммов канал; 5 — угол передней камеры; 6 — радужка; 7 — радужка на срезе; 8—пигментный эпителий радужки
Кольцо Швальбе, как указывалось выше, является передней границей трабекулярной области [980]. Здесь коллагеновые волокна перемешиваются с эластическими волокнами. С воз-
Передняя камера и дренажная система
 растом появляются и спиралевидные коллаге-новые волокна. Кольцо Швальбе является местом перехода эндотелия роговой оболочки к клеткам, покрывающим трабекулы.
растом появляются и спиралевидные коллаге-новые волокна. Кольцо Швальбе является местом перехода эндотелия роговой оболочки к клеткам, покрывающим трабекулы.
Склеральная шпора представляет собой клиновидный гребень, обращенный в сторону полости глаза и состоящий из циркулярным образом ориентированных коллагеновых и эластических волокон (рис. 3.3.4). К склеральной шпоре присоединяется сухожилие продольной ресничной мышцы [615, 910, 959, 980, 1103]. Здесь же присоединяется корнеосклеральная часть трабекулярного аппарата. Переднемеди-альный край шпоры образует задний край склеральной борозды (рис. 3.3.4).
Коллагеновые волокна склеральной шпоры различного диаметра (от 35 до 80 нм). Толщина их увеличивается по мере приближения к склере [1103].
Сокращение ресничной мышцы оттягивает склеральную шпору кзади. При этом открываются межтрабекулярные пространства (рис. 3.3.5). Предполагают, что этот механизм является одним из основных механизмов понижения внутриглазного давления при применении миотиков [411—414, 677—679, 758, 916, 1121].
Недавно было показано, что в пределах склеральной шпоры имеются сократительные миофибробластоподобные клетки, в цитоплазме которых выявлено большое количество а-ак-тинина и миозина [1009, 1060, 1061]. В клетках недостает десмина и микрофилламентов промежуточного типа, т. е. компонентов, характерных гладкомышечным клеткам ресничного тела [1060, 1061]. Миофибробластоподобные клетки склеральной шпоры контактируют с эластическими волокнами склеральной шпоры, а некоторые из них непрерывно переходят в смежные участки трабекулярной сети. Часть клеток трабекулярной сети также содержит а-акти-нин и актин гладких мышц [245, 246, 329]. Таким образом, можно предположить, что сокращение этих клеток может изменять архитектонику трабекулярной сети и изменять сопротивление оттоку камерной влаги.
К некоторым миофибробластоподобным клеткам склеральной шпоры подходят безмя-котные аксоны нейронов, тела которых лежат в супрацилиарном пространстве. Аксоны распространяются в склеральной шпоре циркулярно и параллельно соединительнотканным элементам. Их терминалы плотно контактируют с клеточными мембранами. Окончания нервов содержат зернистый материал и агранулярные пузырьки, напоминающие таковые в адренэргических нервах. Тем не менее волокна не относятся к адренэргическим, что подтверждено иммуногис-тохимически. Tamm et al. [1063, 1064] выявили, что подобного типа пузырьки обнаруживаются в неадренэргических терминалах нервной системы кишечника [393; 394].
Необходимо принять во внимание то, что аксоны, иннервирующие миофибробластоподобные клетки склеральной шпоры у человека, относятся к аминэргическим, пептидэргическим и нитрэргическим. Они в то же время не дают положительной реакции при проведении им-муногистохимической реакции для выявления ацетилхолинэстеразы. Важно отметить и то, что задние участки трабекулярной сети иннер-вируются аналогичным образом [1038, 1064]. Парасимпатические пептидэргические и нитр-эргические волокна, подходящие к склеральной шпоре, исходят из крылонебного ганглия, а также нервных волокон сосудистой оболочки [328, 934].
Предполагают, что пептидэргическая и нит-рэргическая иннервация миофибробластоподоб-ных клеток является основной в регуляции сопротивления оттоку камерной влаги посредством контакта миофибробластоподобных клеток с эластическими волокнами трабекулярной сети. Введение обезьянам нитровазодилята-торов вызывает увеличение оттока камерной влаги [56, 788].
Трабекулярная сеточка(зубчатая связка; reticulum trabeculare; lig. pectinatum; spongium iridocorneale).
На меридианальных срезах глаза видна скудная коллагеновая сеть, выполняющая внутреннюю склеральную борозду и распространяющаяся к корню радужки в виде веера (рис. 3.3.1, 3.3.4). Ручка этого веера располагается несколько кпереди от места прерывания десцеметовой мембраны. Именно в этом месте коллагеновые волокна веера проникают в глубокие периферические слои стромы роговицы и переплетаются с ними.
Трабекулярную сеть можно разделить воображаемой линией на две части. Эту линию необходимо провести от склеральной шпоры к месту прерывания десцеметовой мембраны. Часть трабекулярной сети, лежащую снаружи линии и расположенную между роговой оболочкой и склерой, обозначают роговично-скле-ральной частью (pars corneoscleralis reticulum trabeculare). Часть трабекулярной сети, расположенную кнутри и прилежащую к радужной оболочке, обозначают сосудистооболо-чечной (увеальной) частью (pars uvealis) (рис. 3.3.1, 3.3.4).
Ширина трабекулярной сети сзади, вблизи склеральный шпоры, равняется 120—180 мкм. Она более широкая при близорукости, чем при гиперметропии.
Между корнеосклеральной частью трабекулярной сети и эндотелиальнои выстилкой шлем-мова канала располагается богатая клетками зона — пери- или юкстаканаликулярная соединительная ткань [316].
Пространства радужно-роговичного угла, расположенные между трабекулами (фонта-новы пространства; spatia anguli iridocor-
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 nealis Fontana), содержат гидрофильные гли-козаминогликаны и коллагеновый материал, которые влияют на отток камерной влаги.
nealis Fontana), содержат гидрофильные гли-козаминогликаны и коллагеновый материал, которые влияют на отток камерной влаги.
Сосудистооболочечная (увеальная) часть трабекулярной сети. Внутренняя часть уве-альной трабекулярной сети (1—2 слоя) состоит из переплетающихся трабекул. Самые внутренние трабекулы могут распространяться от ресничной мышцы к кольцу Швальбе. Сзади определяется 2—5 слоев трабекул, внешние слои которых ориентированы циркулярно [65, 66, 154, 1027, 1100ч 1095].
Сзади, трабекулы могут соединяться с циркулярными и радиальными мышечными волокнами ресничной мышцы [83]. Спереди «увеаль-ные трабекулы» постепенно сближаются, и заканчиваются в месте прерывания десцемето-вой мембраны, т.е. внутренней части кольца Швальбе. Эндотелиальное покрытие трабекул постепенно переходит в эндотелий роговой оболочки.
Трабекула увеальной части трабекулярной сети имеет диаметр 4—6 мкм. Она утолщается кзади и сужается кпереди. Ширина меж-трабекулярных пространств колеблется от 20 до 75 мкм.
Роговично-склеральная часть трабекулярной сеточки. Роговично-склеральная часть тра-бекулярного аппарата представляет собой решетчатую уплощенную структуру, состоящую из трабекул. Толщина каждой трабекулы приблизительно 5—12 мкм. Расстояние между тра-бекулами равняется 5—20 мкм. При этом меж-трабекулярные пространства внешних слоев роговично-склеральной части колеблются между 2 и 20 мкм, т. е. пространства более узкие, чем в увеальной части.
Между трабекулами, расположенными на разных уровнях, обнаруживаются межтрабеку-лярные «связки», толщиной от 2 до 5 мкм.
Количество слоев трабекул в роговично-склеральной части колеблется от 8 до 15, а общая ее толщина равна 120—150 мкм. Передние слои роговично-склеральной части тра-бекулярного аппарата сходятся и сливаются с роговичными пластинами [154, 1103].
Трабекула. Основной структурой увеальной и роговично-склеральной частей трабекулярно-го аппарата являются трабекулы [959]. В тра-бекуле различают кортикальную зону и стержень. Снаружи трабекула покрыта одним слоем клеток (рис. 3.3.7—3.3.9).
Клетки трабекулы располагаются вдоль длинной оси трабекулы. Толщина их порядка 4—8 мкм, а длина 120 мкм. Соседние клетки контактируют между собой посредством отростков. Они также соединяются при помощи десмосом и щелевых контактов [877]. Несмотря на наличие межклеточных контактов, радиоактивные трейсеры (ферритин) свободно проникают вглубь трабекулы по межклеточным пространствам.

Рис. 3.3.7. Трехмерное схематическое изображение венозного синуса склеры (шлеммова канала) и трабекулярной сети (по Hogan et al., 1971):
1 — просвет канала; 2 — эндотелиальная клетка; 3 — наружная
стенка канала; 4 — внутренняя стенка канала; 5 — межтрабеку-
лярные пространства; 6 — внутренние соединительные каналы;
7—корнеосклеральные трабекулы
Поверхность трабукулярных клеток покрыта макромолекулами, богатыми сиаловыми кислотными остатками [154, 1090, 1097; 1103]. Между трабекулами гиалуроновый гель не обнаруживается [390].
Трабекулярные клетки содержат обычные органоиды и большое количество пиноцитозных пузырьков [496, 1094]. Обнаруживаются также филаменты цитоскелета. Клетки трабекул отличаются высокой синтетической активностью. Они синтезируют материал базальных мембран, коллаген и гликозаминогликаны [389].
Наиболее важной функцией трабекулярных клеток является их барьерная функция на пути камерной влаги. Эта функция обеспечивается структурными особенностями клеток и зависит от биологической их активности. Одной из функций является также синтез межклеточного материала и его лизис. Последняя функция вытекает из необходимости постоянного лизиса материала, освобождающегося в трабекулярной сети по мере прохождения через нее камерной влаги [410, 653]. О синтетической активности клеток свидетельствуют экспериментальные исследования по культивированию изолированных клеток in vitro. Трабекулярные клетки при этом синтезируют внутри- и внеклеточные гликозаминогликаны (гепарансуль-фат, гиалуроновая кислота, дерматансульфат) [154, 831—835, 903, 912—914, 967].
Получены убедительные данные, свидетельствующие о способности трабекулярных клеток синтезировать волокнистый материал, особенно после травмы или применения кортикостеро-
Передняя камера и дренажная система

|

|
Рис. 3.3.8. Схематическое изображение структурной организации (а) и электронномикроскопическое строение (б)
трабекулы:
/ — эндотелиальная клетка; 2— базальная мембрана; 3— кортикальная зона; 4 — стержень трабекулы
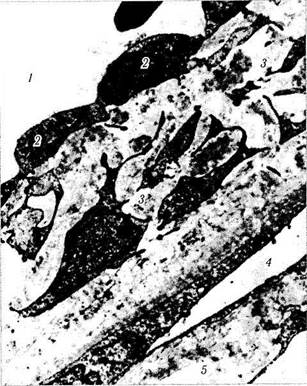
Рис. 3.3.9. Ультраструктурная организация юкстакана-
ликулярной соединительной ткани (по Fine, Yanoff,
1972):
1 — венозный синус склеры (шлеммов канал); 2 — эндотели-
альные клетки, выстилающие шлеммов канал; 3 — юкстакана-
ликулярная сеть; 4— межтрабекулярные пространства; 5 — тра-
бекула
идов [670, 673, 832]. Трабекулярные клетки обладают также фибринолитическими свойствами [814]. В культуре ткани трабекулярные клетки синтезируют в определенном количестве активатор плазминогена.
Трабекулярные клетки обладают высокой фагоцитарной активностью [919]. Нередко в них можно найти зерна пигмента и другие частицы, количество которых увеличивается с возрастом. Введенные в эксперименте частицы (коллоидное золото, пероксидаза хрена, витальные красители) моментально фагоцитируются клетками и, таким образом, выводятся из камерной влаги [94, 194, 541, 919, 1002, 1003]. Для переваривания фагоцитированного материала цитоплазма трабекулярных клеток содержит достаточно большое количество лизосом. У некоторых животных (кошка) после фагоцитоза трабекулярные клетки гибнут и восстанавливаются только спустя 150 дней [541], а у человека поглотившие пигмент клетки сохраняются длительно.
В последние годы установлено, что трабекулярные клетки синтезируют многочисленные биологически активные вещества, некоторые из которых участвуют в регуляции внутриглазного давления. К ним относятся простагландин F2, ингибитор тканевой и матричной металлопро-теиназы. Причем увеличивается синтез этих веществ, и они высвобождаются в камерную влагу при механической деформации клеток, что происходит при колебаниях внутриглазного давления [706, 803].
Интересные данные были получены при изучении синтеза в трабекулярных клетках оксида
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 азота, вещества, обладающего многими функциями. Окись азота обладает иммуномодулиру-ющим свойством, участвует в процессах сокращения и расслабления мышечной ткани, обладает нейромодуляторными свойствами. Образуется окись азота благодаря ферментативной активности синтетазы оксида азота, которая генерирует окисль азота из L-аргинина и является короткоживущим свободным радикалом. Показано, что интенсивность синтеза оксида азота зависит от колебания внутриглазного давления. Колебания давления деформируют трабекулярные клетки, что и является причиной активации синтетазы оксида азота. Такая связь между активацией синтеза оксида азота и обратимой деформацией клеток свойственна не только трабекулярным клеткам. Она характерна для эндотелиальной выстилки шлеммо-ва канала [706, 769, 770], эндотелиальных клеток сосудов [88, 216, 488], хондроцитов [650], остеоцитов [1012].
азота, вещества, обладающего многими функциями. Окись азота обладает иммуномодулиру-ющим свойством, участвует в процессах сокращения и расслабления мышечной ткани, обладает нейромодуляторными свойствами. Образуется окись азота благодаря ферментативной активности синтетазы оксида азота, которая генерирует окисль азота из L-аргинина и является короткоживущим свободным радикалом. Показано, что интенсивность синтеза оксида азота зависит от колебания внутриглазного давления. Колебания давления деформируют трабекулярные клетки, что и является причиной активации синтетазы оксида азота. Такая связь между активацией синтеза оксида азота и обратимой деформацией клеток свойственна не только трабекулярным клеткам. Она характерна для эндотелиальной выстилки шлеммо-ва канала [706, 769, 770], эндотелиальных клеток сосудов [88, 216, 488], хондроцитов [650], остеоцитов [1012].
Оксид азота способен расслаблять трабе-кулярную сеть и ресничную мышцу [1169]. Происходит это благодаря существованию различных механизмов. Так, оксид азота, синтезируемый трабекулярными клетками, может включать гуанилил циклазу и различные ауто-кринные и паракринные механизмы, приводя к увеличению концентрации циклического GMP в трабекулярных клетках [140, 753]. О роли оксида азота, синтезируемого трабекулярными клетками, свидетельствуют наблюдения снижения активности синтетазы оксида азота при глаукоме [769, 770].
Непосредственным механизмом влияния оксида азота на регуляцию внутриглазного давления является его влияние на концентрацию в цитоплазме трабекулярных клеток ионов кальция. Показано, что в трабекулярных клетках при повышении ВГД до 20—30 мм Hg изменяется концентрация внутриклеточного кальция [707], поскольку синтетаза оксида азота (bNOS и eNOS) активизирует комплекс каль-ций/кальмодулин (комплекс кальция с кальмо-дулином является месседжером, изменяющим активность многих ферментов, регулирующих кальциевый насос, различные специфические белковые киназы, циклические нуклеотидные фосфодиэстеразы, гистоны и тубулин) [140, 753].
Кортикальная зона. Кортикальная зона состоит из окрашивающегося положительно ШИФФ-реактивом пластинчатого материала, присоединенного к трабекулярным клеткам при помощи полудесмосом. Внутренняя граница этой зоны не очень хорошо видна и инфильтрирована соединительнотканными элементами коры.
В пределах базальной пластинки найдены скопления веретенообразных коллагеновых волокон с периодичностью, колеблющейся от 30—40 до 80—120 нм [387, 697, 1070, 1103].
Стержень. Стержень каждой трабекулы образован коллагеном I, II и IV типов. Он также содержит фибронектин, эластин, хондро-итинсульфат, дерматансульфат и спиралевидный коллаген [326, 387—389, 698, 909, 1070, 1092, 1103].
Коллагеновые фибриллы (толщина 30— 50 нм) ориентируются вдоль длинной оси трабекул. В трабекулах увеальной части они формируют компактный стержень. Ориентация коллагеновых фибрилл в трабекулах, вероятно, определена направлением приложения силы при сокращении мышц ресничного тела.
В «увеальной» трабекуле эластические волокна располагаются, главным образом, в центре стержня. Эти эластические волокна отличаются по строению от эластических волокон других тканей организма. Состоят они из волокнистого и аморфного компонентов [1103]. В этой области иммуногистохимически определяется большое количество микрофибриллярного белка, близкого к эластину,— фибрил-лина [1162]. Ультраструктурно показано, что только центральная зона эластического волокна содержит эластин и тропоэластин. Эти компоненты погружены в электронноплотный материал неизвестной природы [388, 671].
Эластический компонент трабекулы играет определенную роль в способности трабекулы к сокращению, что было показано на изолированной трабекуле быка [654].
Сокращению способствует наличие в трабекулярных клетках миофиламентов (актин). Именно эта особенность позволяет отнести трабекулярные клетки к миофибробластам [329]. Показано, что у человека количество таких клеток уменьшается с возрастом. Сохраняются они лишь вблизи склеральной шпоры [1061].
Особого внимания заслуживают вопросы возрастных изменений трабекулярных клеток. С возрастом пролиферативная активность трабекулярных клеток снижается [968]. Кроме того, на протяжении жизни количество клеток постоянно линейно уменьшается со скоростью потери 0,56% клеток в год [61]. Количество клеток у 20-летнего индивидуума равняется примерно 763 000, а у 80-летнего — всего лишь 403 000. При этом количество клеток уменьшается ежегодно примерно на 6000 [404]. Интересно, что скорость потери трабекулярных клеток различна в различных участках тра-бекулярной сети. Наибольшая потеря клеток отмечается в центральных участках [60, 61, 404, 416].
Немаловажное практическое значение имеет выявление репаративных возможностей трабе-кулярной сети. Трабекулярные клетки in vitro не способны регенерировать. Тем не менее при повреждении трабекулярной ткани отмечаются признаки регенерации клеток, принимающих кубовидную форму. При этом увеличивается их число. Подобную регенерацию трабекулярных
Передняя камера и дренажная система

 клеток выявляли после трабекулоэктомии или после лазерной трабекулопластики [41].
клеток выявляли после трабекулоэктомии или после лазерной трабекулопластики [41].
Отростки радужной оболочки представляют собой однородные треугольной формы «связки», идущие от корня радужки до трабе-кул «увеальной» части трабекулярного аппарата, с которыми они и сливаются. Иногда отростки достигают склеральной шпоры, а иногда и линии Швальбе. Количество их незначительно. Обнаруживаются они у трети индивидуумов. У индивидуумов с карими глазами эти отростки пигментированы. Строение отростков аналогично строению стромы радужки. Иногда отростки прикрывают угол передней камеры.
Клетки Швальбе. В месте перехода между роговой оболочкой и трабекулярной сетью рядом исследователей обнаружены клетки, отличающиеся хорошо выраженной эндоплазма-тической сетью, большим количеством митохондрий и многочисленных электронноплотных гранул. Эти клетки были названы клетками Швальбе [873]. Предполагают, что клетки Швальбе обладают секреторной активностью, о чем свидетельствует не только обнаружение в цитоплазме гранул, дающих положительную реакцию при выявлении нейрон-зависимой эно-лазы, гиулоронат-синтетазы [1039]. Происхождение и функция этих клеток пока неизвестны.
Пери- или юкстаканаликулярная соединительная ткань распространяется вдоль всего шлеммова канала (рис. 3.3.9). Толщина этой зоны колеблется от 2,0 до 20,0 мкм, и располагается она между эндотелиальной выстилкой канала и лежащей кнутри «корнеосклеральнои» частью трабекулярного аппарата. Эта зона складывается из 2—5 клеточных слоев, погруженных в межклеточное вещество (рис. 3.3.9). Клетки обладают длинными отростками и соединяются между собой при помощи зон замыкания, десмосом и щелевых контактов. Между клетками определяются промежутки шириной 10 мкм, через которые проникает камерная влага по направлению эндотелиальной выстилки шлеммова канала [1103]. Между этими клетками и эндотелием шлеммова канала располагается базальная мембрана.
Периканаликулярные клетки обладают важными функциями — фагоцитарной и синтетической. Эта ткань представляет собой наиболее мощное препятствие на пути оттекающей влаги передней камеры глаза. Связано это не только с тем, что межклеточные пространства узкие и извилистые, но, в первую очередь, с присутствием внеклеточно расположенных протеогли-канов и гликопротеидов [114, 297, 409, 521, 670, 671, 992, 1132].
Зона, контактирующая с внешней стенкой шлеммова канала, содержит меньше клеток, чем прилегающая к ней трабекулярная ткань. Состоит она из 4—8 плотно упакованных слоев фиброцитоподобных клеток. Толщина этой зоны порядка 5—15 мкм. Помимо клеток, в ней
определяются неравномерно распределенные коллагеновые, эластические волокна и мелкозернистый материал. Коллаген относится к VI типу [675].
Имеется также и переходная зона, толщиной 20—30 мкм, располагающаяся между этой юкстаканаликулярной тканью и склерой. Она состоит примерно из 10 коллагеновых пластин, практически идентичных склеральным пластинам.
Межклеточное вещество. Главными компонентами межклеточного вещества являются коллаген I, III,IV, V и VI типов, фибронек-тин, хондроитин- и дерматансульфат. Обнаруживается также гиалуроновая кислота и эластическая ткань. Многие из этих макромолекул (коллаген VI типа, фибронектин, хондроитин- и дерматансульфат) содержат сиаловую кислоту. Обнаружен и фибриллин [1162]. Особенностью межклеточного вещества является наличие эластических волокон, образующих густую объемную сеть («решетчатое сплетение») [907]. Поскольку эластические волокна связаны с сухожилиями мышцы ресничного тела и базаль-ной мембраной эндотелиальных клеток шлеммова канала, они могут влиять на проходимость этой области для камерной влаги [388, 389, 671, 680, 907].
Дата добавления: 2015-03-26; просмотров: 1188;