Основные узлы и механизмы станка.
Понятие «эволюция» в биологию было введено швейцарским ученым Ш. Боннэ (1762). Он понимал под этим термином не только саму идею развития, но и отмечал изменчивость и некий отбор в становлении форм живого. Эволюция – неотъемлемое свойство живой природы. Идеи единства и развития природы можно проследить с древнейших времен. Проблемы происхождения и эволюции жизни на Земле были в центре философских и религиозных систем. Так, Г.Лейбниц, развивая идеи Аристотеля, провозгласил принцип градации и предсказал существование переходных форм между растениями и животными. В России эти идеи активно поддерживал А.Н. Радищев. В дальнейшем этот принцип был развит в представлении о «лестнице существ» от минералов до человека и Бога, доказывая трансформизм живой природы. Так выявлялась не эволюция, а общность и сходство форм и усложнения организмов. Но сопоставлялись время существования Земли и время формирования форм живого на ней. Бюффон обосновывал в своей «Естественной истории» историю Земли и доказывал единство происхождения живого планом строения. Другой энциклопедист, Д.Дидро, считал, что разнообразие органического мира объясняют мелкие изменения живых существ и длительность существования Земли. У Мопертьюи были догадки о корпускулярной природе наследственности, эволюционной роли вымирания неприспособленных форм и роли изоляции в возникновении новых форм. Кант в «Космогонии» говорил о развитии живого мира в течение миллионов лет.
К. Линней считал, что близкие виды внутри рода могли развиться естественным образом без участия высших сил. Эволюционные представления были характерны для К.Вольфа, М.В.Ломоносова и А. Н. Радищева. Ломоносов считал изменения в неживой природе причиной изменений мира живого, по останкам вымерших форм судил об условиях существования их в далекие времена. Он писал, что время, необходимое для создания организмов, больше, чем определяется церковным исчислением. Но эти идеи формировали пока только представление о последовательности природных тел. Ограниченную трансформацию видов допускал Ж. Бюффон, считая, что разные типы животных имеют разное происхождение и возникли в разное время. Эта концепция обобщала многие наблюдения и факты, выделяла идею глубокой взаимосвязи между видами, подвидами, родами и другими таксонами, подготавливая почву для эволюционизма. Так, до конца XVIII в. господствовала мысль о «целесообразности порядков в природе» (сотворении кошек для пожирания мышей и т.п.). Постепенно возникал вопрос о возникновении такой целесообразности. Общество не могло еще воспринять идеи эволюции, так как обсуждались не доступные для проверки масштабы времен.
Идею эволюции живого перевел на уровень теории эволюции Ж.-Б. Ламарк.Он считал, что Бог сотворил материю и движение, а далее развитие происходило по естественным причинам. Опираясь на многочисленные факты изменяемости видов, Ламарк в книге «Философия зоологии» (1809) выдвинул гипотезу о механизме эволюции, основанном на двух предпосылках: наследование приобретенных признаков и упражнение или неупражнение частей организма. Он представил эволюционное обоснование «лестницы существ», основанное на принципах градации (внутреннего стремления к совершенству) и изначальной целесообразности реакции организма на изменения внешней среды (признание возможности прямого приспособления). Далее Ламарк формулировал два закона: 1 — изменение привычек следует сразу за изменением условий и 2 — эти изменения передаются по наследству. Основа эволюции — врожденная способность к самосовершенствованию, фактор явно нематериальный: «творить может только Бог, тогда как природа может только производить», а изменения во внешней среде могут изменить формы поведения, поэтому органы или структуры способны приобрести новые функции, а эти новые функции органов и изменения в них могут быть переданы потомкам. Так вытянулась шея у жирафа, увеличились перепонки у водоплавающих, развивается мускулатура при занятиях спортом. Эта часть учения Ламарка отвергалась, как противоречащая появляющимся новым знаниям о механизме наследования, для нее еще не пришло время. Большую роль в возникновении новых видов Ламарк отводил переменам климата и гидрогеологического режима. Политические страсти внутри биологии скомпрометировали важность идей типа «наследуется все благоприобретенное» (она получила название «ламаркизм»). Как подчеркивал К.А.Тимирязев, Ламарк не сумел объяснить целесообразность организмов.
Учение катастрофизма отражало идеи развития природы, сформировавшейся к началу XIX в. Французский зоолог Ж. Кювье выделял четыре типа животных — позвоночные, мягкотелые, членистые и лучистые. И с каждым из них он сопоставлял некий «план композиции», некую «творящую силу», которая после очередной катастрофы в геологической истории обеспечивает восхождение органических форм. Тем самым геологическую эволюцию планеты Кювье связывал с эволюцией живого, признавая роль катастроф и неравномерности темпов преобразований в природе, что не потеряло своего значения и поныне. Ему возражал У. Смит, один из основоположников биостратиграфии, отмечавший непрерывность распространения сходных видов в близких по возрасту слоях.
Концепцию униформизма сформировали противники теории катастроф. Дж. Геттон, Ч. Лайель, М.В. Ломоносов критиковали идеи Кювье за неопределенность причин катастроф, за укорачивание возраста Земли. Под впечатлением успехов классической механики они считали мир познаваемым и предлагали опираться на преемственность настоящего и прошлого, выделяя непрерывность действия законов и факторов в истории Земли и возможность обратимости явлений. При эволюции живого Лайель допускал возможность актов творения, демонстрируя прогрессивные изменения ископаемых останков. Униформисты считали, что вымирание несовместимо с естественным образованием новых видов, и потому предполагали участие творца. Фактически они свели историю планеты к цикличности и случайным изменениям.
Униформизм, как и ламаркизм и катастрофизм, предварял теорию естественного отбора Дарвина, частично конкретизировал идею эволюции. Английский экономист Т.Мальтус привлек внимание к репродуктивному потенциалу человека и указал на экспоненциальный рост численности населения. В 1788 г. он опубликовал «Трактат о народонаселении», в котором убедительно и ярко обрисовал, к чему может привести ничем не сдерживаемый рост населения. Благодаря Дарвину выражение Мальтуса «борьба за существование» приобрело широкую известность. Они оба считали ее результатом несоответствия между быстрым ростом популяций и ограниченностью пищевых ресурсов. Идеи Мальтуса и Лайеля оказали большое влияние на Ч.Дарвина. Он верил в познаваемость законов природы, в возможности объяснения наблюдений. Дарвин считал проблему происхождения человека связанной с эволюцией неорганического и органического мира. К середине XIX в. в разных областях биологии был накоплен огромный фактический материал, который нуждался в обобщении. Да и практика сельского хозяйства требовала теории, которая бы открыла пути селекции.
Наконец, Ч. Дарвин опубликовал в 1859 году знаменитый труд «Происхождение видов путем естественного отбора», создав теорию эволюции, обобщив отдельные эволюционные идеи и разрешив накопившиеся противоречия. У него эволюция определяется триадой: наследственность, изменчивость и естественный отбор.
Основой теории эволюции путем естественного отбора послужил огромный материал, собранный и до Дарвина, и им самим. Он предпринял пятилетнее морское путешествие на корабле «Бигль», во время которого занимался геологическими исследованиями, собирал ботанические, зоологические и палеонтологические коллекции. Кульминацией, с точки зрения формирования его эволюционных взглядов, явилось исследование флоры и фауны Галапагосских островов, где он увидел в действии процесс эволюции при сравнении близких видов вьюрков, ящериц, черепах. По прибытии в Англию он проанализировал историю селекции и выявил отличия между породами и сортами. Здесь он усмотрел творческое начало в деятельности селекционеров, позволяющее накопить изменения в результате отбора.
Дарвин собрал обширный материал об изменчивости организмов и видов и отметил почти всегда постоянную численность популяций. Способность к размножению, свойственная всему живому, обеспечивает сохранение вида. Численность популяций на Земле контролируется различными факторами среды (пространство, свет, пища, тепло). Исследуя и сопоставляя огромный материал и находясь под впечатлением идеи Мальтуса, Дарвин начал понимать, что при интенсивной конкуренции между членами популяции любые изменения, благоприятные для выживания в данных условиях, повышают способность особи к размножению и оставлению плодовитого потомства. Но каждый вид производит больше особей, чем выживает их до взрослого состояния, а среднее число взрослых особей почти постоянно. Ненужные формы при этом отбрасываются путем нового механизма — естественного отбора. Черновой вариант своей теории он сделал в 1842 г.
Понятиям изменчивость и наследственность, которые Ламарк связывал с приспособляемостью, передаваемой по наследству и являющейся основой видообразования, Дарвин придал принципиальное значение. Определенная изменчивость — это способность всех особей определенного вида одинаковым образом реагировать на изменения среды, при этом изменения в организмах не наследуются (сейчас это — адаптивная модификация). Неопределенная изменчивость приводит к существенным изменениям в организме, которые наследуются с усилением в следующих поколениях (мутация, по современной терминологии). Она тоже связана с условиями окружающей среды, но не непосредственно. Дарвин считал, что именно такая изменчивость играет ведущую роль в эволюции.
Естественный отбор, или выживание наиболее приспособленных — механизм эволюции, материал для него — наследственная изменчивость. В нем Дарвин соединил многие биологические знания, в том числе опыт практической селекции.
Так пришли к представлению о том, что органический мир представляет некое единство, имеет свою историю, а его нынешнее состояние есть результат предшествующего. Заслуга Дарвина в том, что из сопоставления фактов борьбы за существование и всеобщей изменчивости свойств и признаков он вывел неизбежность избирательного уничтожения одних особей и размножения других — естественного отбора. Начинаясь с наблюдения, познание жизни продолжалось на уровне мыслительных процессов. В классической биологии эксперимент еще не был методом познания живого. Механистический детерминизм игнорировал функциональное единство живых систем, а телеологический подход основывался на целесообразности организмов. С теории эволюции Дарвина, в основе которой лежал рациональный подход, началось преодоление идеалистической тенденции в биологии.
Учение Дарвина существенно потеснило антиэволюционные и креацианистские концепции. При этом сопоставляли данные палеонтологии, сравнительной анатомии и эмбриологии (метод Геккеля). Данные палеонтологии доказывали существование эволюции живого. Это показал еще В.О. Ковалевский на примере развития вида лошадей, обнаружив существование предка с пятипалой конечностью, жившего 60 млн лет назад. Последовательные ряды ископаемых животных он выстроил в ряд для наглядности эволюционных изменений. (Такие ряды называют филогенетическими.) До конца XIX в. эволюционные идеи овладевали умами, строились филогенетические древа для всех крупных групп растений и животных. Существуют и эмбриологические доказательства эволюции. Закон Геккеля — Мюллера утверждает, что каждая особь в своем индивидуальном развитии (онтогенезе) повторяет историю развития своего вида (филогенез). К доказательствам относят и наличие рудиментальных органов, и явления атавизма. Существуют и биогеографические доказательства: сравнение животного и растительного мира разных континентов показывает, что различия внутри вида тем больше, чем дольше длилась их изоляция. Биоразнообразие — неизбежный результат отбора.
В результате изучения эволюции надвидовых групп (макроэволюции) сформировался так называемый классический дарвинизм: установлена необратимость эволюции, принцип мультифункциональности органов и эволюции органов путем смены функции, биогенетический закон и др. Но многие ученые не приняли дарвинизм, и это неприятие и критика особенно усилились в период возникновения генетики. Можно даже сказать, что распространение эволюционных идей сопровождалось в это время острой критикой теории естественного отбора. Отсутствие обнаружения переходных форм препятствовало принятию дарвиновских идей, а с появлением генетики естественный отбор все больше подвергался критике. Так, Ф. Дженкин показывал, что при скрещивании произойдет «растворение признаков», которые были единично поддержаны отбором. Основу такой критики составило отсутствие строгих доказательств наследственной изменчивости, которые давала генетика (теория мутаций, учение о чистых линиях и принцип корпускулярной наследственности). В.Л. Иоганнсен показал неэффективность отбора в чистых линиях (в потомстве одной самооплодотворяющейся особи). Г.де Фриз выдвинул мутационную гипотезу о скачкообразном возникновении новых видов путем крупных изменений наследственности (мутации) без ведущего участия естественного отбора.
Постепенно стал складываться синтез генетики и классического дарвинизма – синтетическая теория эволюции (СТЭ). Выявлялись новые факторы эволюции, уточнялась терминология. После опытов Вейсмана и появления мутационной гипотезы де Фриза были вскрыты закономерности распределения хромосом при клеточном делении.
Хромосомная теория наследственности, сформулированная А. Вейсманом, выдвинула принцип невозможности передачи по наследству «благоприобретенных» признаков. Отрезанные хвосты у мышей во многих поколениях даже не укорачивались. Приобретенные признаки не влияли на половые клетки, передающие признаки следующим поколениям, роль среды фактически сводилась к сортированию возникающих независимо от нее наследственных изменений. Требовалось уточнение понятия изменчивости. Т. Морган установил, что признаки, гены которых «сцеплены» в одной хромосоме, наследуются совместно. Поэтому и третий закон Менделя выполнялся не всегда. После обнаружения у дрозофилы групп сцепления генов по числу имеющихся хромосом эта теория оформилась (Т. Г. Морган, А. Стертевант и др.). В 1950 г. была найдена тонкая структура гена (С. Бензер), понят язык, на котором была записана генетическая информация. Эти генетические механизмы наследственности существенны и для понимания изменчивости как основы отбора.
Изменчивость — способность живых организмов приобретать новые признаки и свойства, отражающая взаимодействие организма с внешней средой. Различают наследственную (генотипическую, или мутационную) изменчивость и ненаследственную (модификационную) изменчивость (вместо неопределенной и определенной изменчивости у Дарвина соответственно). Первая связана с мутациями, возникает из-за изменения структуры гена или хромосом и служит единственным источником генетического разнообразия внутри вида. Причиной мутаций могут быть внешние жесткие излучения, химические причины и прочие мутагены (например, вирусы). Большая часть мутаций рецессивна и не проявляется у гетерозигот. Рекомбинации при половом размножении также порождают множественные мутации, которые приводят к комбинативной изменчивости, создающей материал для естественного отбора. Но новых видов при этом не образуется. При изменении внешних условий некоторые ранее вредные рецессивные мутации могут оказаться полезными, и их носители могут получить преимущество при естественном отборе. Мутации — фактор случайный, подчиняющийся статистическим законам. Поэтому они, как и перестройки генов, и волны численности популяции, не могут быть решающим фактором эволюции.
Модификационная изменчивость — это сходные изменения признаков у всех особей потомства популяции какого-то вида в сходных условиях существования. Она не затрагивает гены и не передается по наследству. Модификационные изменения адаптационны, т. е. делают популяцию лучше приспособленной к изменению условий. Они могут происходить лишь в пределах генотипа и не выходят за пределы нормы данного признака и для эволюции не существенны. Благодаря изучению генетических процессов в популяции эволюционная теория интенсивно развивалась.
Генные мутации — главная причина возникновения новых наследственных свойств. Они и есть основные предпосылки эволюции, постоянно действующий источник наследственной изменчивости. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и природные популяции насыщены самыми разнообразными мутациями из-за одновременных мутаций многих генов.
Рецессивные мутации могут накапливаться в генофондах популяций, составляя резерв наследственной изменчивости. Классические работы С.С. Четверикова связали закономерности отбора в популяциях с динамикой процесса эволюции. На нескольких видах мушек дрозофил он показал, что в каждой популяции есть большое количество разных рецессивных мутантных генов. Эти гены не выявляются в признаках организма, так как подавлены нормальными доминантными аллелями, но могут проявиться в случае, когда встретятся и оставят потомство две особи с одним и тем же рецессивным мутантным геном. Так колебания частоты генов в популяциях связаны с внешними условиями среды.
Этот закон Четверикова был многократно проверен и на других объектах, породив мнение, что наличие таких рецессивных мутантных генов является предпосылкой эволюции. Но важно, чтобы эти особи не имели дефектов, мешающих нормальной работе организма, и обладали какими-то преимуществами перед другими. Так, при близкородственном скрещивании (инбридинг) потомство оказывается гомозиготным не только по этому мутантному гену, но и по большим отрезкам хромосом, что не способствует эволюции.
Основную роль должны играть доминантные мутации, считает Гершенсон, а рецессивные — могут изредка поддерживаться отбором, хотя у дрозофил численность рецессивных мутантных генов велика, но частота каждого мала (порядка сотых долей процента). Такой случайный спектр рецессивных мутаций свидетельствует о ненаправленном характере мутационного процесса в популяции. Доминантные же мутации почти все принадлежат к очень ограниченному числу типов и вызывают лишь несколько определенных небольших изменений в структуре жилок крыльев, числе и расположении щетинок. В популяциях их доля около 15 %, сохраняющаяся из года в год. Отсюда и предположение о действии естественного отбора, обеспечивающего и сохранность типов.
Мутации как бы нащупывают экологические условия, способствующие выживанию и размножению особей с данной мутацией. Одновременно идет отбор генотипов, в которых она наиболее благоприятна. Важно ее влияние и на норму реакции организма. Так, мутантный признак закрепляется в наиболее подходящих местах, где мутанты становятся постоянной частью природной популяции. Затем на стадии сосуществования мутантов с немутантами происходит приспособление популяции к более эффективному использованию среды обитания. При этом эволюционная пластичность популяции высока и позволяет быстро перестроиться при стойких изменениях среды. Появившаяся мутация может повысить адаптивные свойства организма, тогда можно говорить о третьем этапе, о появлении нового экотипа. И если какой-нибудь из этих экотипов окажется в изоляции от других популяций вида, то начнет образовываться новая разновидность, способная стать и новым видом. Эта схема сильно упрощена, не учтено множество факторов, могущих повлиять на процесс, в том числе и недавно открытых, — перемещения гена в пределах генома, умножение числа какого-то гена в геноме и т. п.
Закон Харди — Вайнберга — без внешних давлений частоты генов в популяции постоянны (1908) — служил первым существенным шагом к объединению дарвинизма и генетики. Этот закон означает, что накопленные изменения в генофонде не исчезают бесследно. Исходя из него и учитывая влияние отбора и возникновение новых мутаций, С.С. Четвериков показал, что из-за постоянных мутаций во всех популяциях создается существенная наследственная гетерогенность, что отбору подвергаются не отдельные особи и виды, а генотип популяции. С работы С.С. Четверикова «О некоторых моментах эволюционного процесса с точки зрения современной генетики» (1926) наступил период синтеза представлений. Мутации — основа эволюции, они перерабатываются естественным отбором. Исследования конца 20-х гг. XX в. показали, что большую роль в эволюции играет не только появление новых мутаций, но и изменение частоты встречаемости существующих аллелей (гена) из-за случайных процессов — колебания численности популяций и пр. (Р.А. Фишер, Н. П.Дубинин, Д.Д. Ромашов, С.Райт и др.). При резком снижении численности популяций (в связи с ростом близкородственных скрещиваний) снижается наследственная изменчивость. По Райту — это «дрейф генов», а по Дубинину — «генетико-автоматический процесс». Другим проявлением «волн жизни» является изменение концентрации различных мутаций и уменьшение разнообразия генотипов популяции. Они могут привести к изменениям направленности и интенсивности действия отбора.
Изоляция популяций необходима для образования нового вида, она — также важнейший фактор микроэволюции. Изоляция как фактор видообразования может достигаться различным образом:
1. Географическая изоляция связана с расширением зоны обитания (ареала). В новых условиях постоянно происходят мутации, наследственные изменения, действует естественный отбор, что приводит к новому виду. Преградами могут быть реки, горы, ледники и пр. Образование вида таким образом занимает сотни и тысячи поколений. Дарвин выделял роль среды в видообразовании. Животные, обитающие на островах Зеленого Мыса, несмотря на некоторое сходство с материковыми видами, имели существенные различия.
2. Временная изоляция достигается несовпадением сроков размножения между двумя подвидами. В результате подвиды расходятся еще больше и возникают два новых вида. Таких примеров много среди рыб.
3. Репродуктивная изоляция возникает из-за различий в поведении или несовместимости генетического материала.
Генетика позволила проследить протекание эволюционного процесса от появления первого признака в популяции до возникновения нового вида. При исследованиях на микроэволюционном(внутривидовом) уровне применялись точные экспериментальные методы. И пришли к элементарной единице эволюции — популяции, элементарном эволюционном материале и явлении. Учение о микроэволюции сформулировали Ф.Г.Добжанский и Н.В.Тимофеев-Ресовский (1939). Современная теория не только добавила к дарвиновской «триаде» новые факторы эволюции, но и основные факторы переосмыслила иначе. Сейчас к ведущим факторам эволюции относят мутации, популяционные волны численности и изоляцию. Возникла и глобальная цель — управление процессом эволюции.
Популяция генетически обособлена от других популяций того же вида и обладает общим генофондом, что обеспечивает генотипическое сходство входящих в нее особей. Из-за малой продолжительности жизни отдельной особи по сравнению с временами эволюции ее генотип на эволюции не скажется. Возникшие наследственные изменения особи в силу свободного скрещивания могут распространиться в популяции, создавая генетическую неоднородность особей и условия для отбора. Популяция — часть вида, т. е. входящие в нее особи принадлежат к одному виду (генетически замкнутой системы, представители которой не могут скрещиваться и давать плодовитое потомство с представителями других видов). Поэтому возникшая мутация не выйдет за пределы вида, и реальные эволюционные сдвиги можно обнаружить лишь в популяциях. Значит, популяция — элементарная биологическая единица, в которой возникают эволюционные процессы.
Синтетическая теория эволюции появилась в 30 — 40-е гг. XX в., объединив разные учения на основе дарвинизма, данных генетики и экологии. В ней популяцию признают в качестве основной единицы эволюции и выделяют два типа эволюции — на микро- и макроуровнях.
Микроэволюцию составили несколько разделов биологии. Среди них — генетико-экологическое изучение структуры популяции (Н.И.Вавилов, Е.Н.Синская, Дж.Клаузен, М.А.Розанова), экспериментальное и теоретическое изучение борьбы за существование и естественного отбора (В. Н. Сукачев, Дж. В. Холдтейн, Г. Ф. Гаузе и др.), данные теоретической и экспериментальной генетики (М.Лернер, И.И.Шмальгаузен, Н.П.Дубинин, Г.Стеббинсидр.), развитие теории вида (Н.И.Вавилов, Э.Майр, К.М.Завадский и др.). Теория микроэволюции изучает необратимые преобразования генетико-экологической структуры популяции, которые могут привести к образованию нового вида.
Теория макроэволюции сформировалась в работах Н.И.Вавилова, И.И.Шмальгаузена, Дж. Г. Симпсона, А.Н.Северцева и др. Она изучает происхождение надвидовых таксонов (семейств, отрядов, классов и пр.), основные направления и закономерности развития жизни на Земле в целом. Эти процессы недоступны наблюдению и могут быть только реконструированы. Всех этих исследователей можно назвать творцами «второго синтеза» (после первого, осуществленного Дарвином) биологического знания – СТЭ.
Основные положения синтетической теории эволюции (неодарвинизма) таковы:
1. Естественный отбор — главный движущий фактор эволюции, является следствием конкурентной борьбы за существование, особенно острой внутри вида и популяции. Факторами образования видов являются мутации, дрейф генов и различные формы изоляции.
2. Расхождение признаков организмов в ходе эволюции от общего предка (дивергенция) происходит через отбор мелких случайных мутаций. Новые формы образуются через крупные наследственные изменения, жизненность которых определяет отбор.
3. Исходным материалом эволюции служат мутации, случайные и ненаправленные. Организация популяции и изменения условий среды выделяют наследственные изменения в сторону прогресса.
4. Макроэволюция, ведущая к образованию надвидовых групп, осуществляется через процессы микроэволюции. Макроэволюция — процесс формирования более крупных единиц: из видов — новых родов, из родов — новых семейств и т.д. Эти процессы нельзя изучать непосредственно, поскольку они очень длительны. Но в основе макроэволюции лежат те же движущие силы, что и в микроэволюции: наследственная изменчивость и начало дивергенции; естественный отбор и продолжение дивергенции, гибель менее приспособленных и образование новой структурной единицы; репродуктивное разобщение, что доказывается несколькими независимыми путями:
Положение об элементарных явлениях и факторах эволюции сформулировал Н.В.Тимофеев-Ресовский: а) популяция — элементарная эволюционная структура; б) изменение генотипа популяции — элементарное эволюционное явление; в) генофонд популяции — элементарный эволюционный материал; г) элементарные эволюционные факторы — мутации, «волны жизни», изоляция, естественный отбор. Отбор проявляется в трех формах: движущей (приводящей к сдвигу видовой нормы), дизруптивной («разрывающей» популяцию на части из-за элиминации прежней нормы), стабилизирующей (поддерживающей существующую норму). Движущий отбор проявляется при изменении условий существования вида. Его давление направлено в пользу особей, имеющих отклонение определенного признака от нормы. Происходит сдвиг общей нормы и возникает новая. Дивергенция между старой и новой нормами ведет к видообразованию. Движущий отбор лежит в основе появления популяций насекомых, устойчивых к определенному яду. Эти особи приобретают преимущества при размножении, и их потомки занимают места умерших насекомых, которые не обладали этим признаком. Таким путем исчезли и многие органы, не используемые несколькими сотнями поколений.
Разрывающий отбор действует при изменении условий существования, его давление направлено в пользу организмов, имеющих отклонения от нормы в обе стороны. И формируется новая норма реакции. Так, на островах, где сильны ветры, мухи с нормальными крыльями сдуваются и гибнут. Преимущество у мух или с недоразвитыми крыльями (они ползают), или с длинными крыльями (они хорошо летают и оказывают сопротивление ветру).Стабилизирующий отбор действует в почти неизменных условиях существования. Он оказывает давление в пользу особей, имеющих средние значения какого-то признака. В результате происходит их укрепление, предохранение от разрушающего действия мутаций. И в местностях, где условия жизни не менялись, сохранились древние виды, вымершие в других местах. Например, так сохранились голосеменное растение гингко, кистеперая рыба латимерия.
Биологический прогресс — результат успеха в борьбе за существование. Он характеризуется возрастанием численности особей, расширением ареала обитания, увеличением числа групп более низкого ранга. Биологический регресс характеризуется обратными признаками и ведет к вымиранию. К биологическому прогрессу ведут следующие факторы:
морфологический прогресс — усложнение организма, поднятие его на более высокий уровень. Строение организма изменяется не вследствие приспособления к изменяющимся условиям среды, оно позволяет расширить использование условий внешней среды. При дальнейшей эволюции эти изменения, называемые арогенезом (совокупностью ароморфозов), сохраняются и ведут к возникновению новых групп, видов;
аллогенез — эволюционное направление, сопровождающееся идиоадаптацией — приспособлением к специальным условиям среды, полезным в борьбе за существование, но не меняющим уровня организации. Пример — колючки растений или изменение окраски животных;
катагенез — эволюционное направление, сопровождающееся упрощением организации. Фактически — это морфологический регресс. Пример — переход к паразитическому образу жизни, уменьшающий способности к конкурентной борьбе.
После каждого «шага» морфологического прогресса начинается приспособление отдельных популяций к условиям существования путем идиоадаптации. Например, класс птиц при расселении по суше дал огромное разнообразие форм. Хотя основы их строения одинаковы, частные приспособления отличны. Поэтому чередование этих главных направлений отражает эволюционную тенденцию в филогенезе почти всех групп.
В рамках современной «классической» (дарвиновской) парадигмы биологии изучение проблемы эволюционного прогресса выступает в качестве одной из основных задач; она получила достаточно глубокую, хотя (по общему признанию) и не исчерпывающую разработку. В основном эта разработка лежит в русле изучения главных направлений эволюционного процесса.
Ещё в 1925 году известный отечественный дарвинист Алексей Николаевич Северцов предложил понимать биологический прогресс данной группы организмов как процесс, связанный с численным увеличением и расселением этой группы, а также с дифференциацией её на подчинённые систематические единицы (например, вида - на подвиды, отряда-на подотряды и семейства и т.д.); такой биологический прогресс представляет собой ничто иное, как победу представителей данной группы в борьбе за существование. При этом Северцов выделял несколько «частных случаев» или способов достижения биологического прогресса-в первую очередь речь идёт об 1) ароморфозе (или морфофизиологическом прогрессе), связанном с ростом «общей энергии жизнедеятельности», общего уровня организаций, (биологическое понятие для обозначения уровня интенсивности, сложности и разнообразия функций органов данного живого существа); иными словами, ароморфоз-это усложнение, 2) идиоадаптации - направлении эволюционного процесса, не связанном ни с повышением ни с понижением «общей энергии жизнедеятельности, при сохранении неизменности общего уровня организации; 3) общей дегенерации-направлении эволюционного процесса, связанном со снижением общей энергии жизнедеятельности и общего уровня организации.
К ароморфным изменениям относятся, например, изменения в ряду: прокариоты- эукариоты- многоклеточные; (в частности - хордовые), а среди позвоночных - в ряду: рыбы-амфибии-рептилии-птицы и млекопитающие (основные классы типа хордовых).
Идиоадаптации связаны с возникновением в эволюции более частных изменений и широкой адаптивной радиацией в рамках уже достигнутого уровня организации. Идиоадаптации рассматриваются как результат предшествующего ароморфоза и, в принципе, развитие по пути идиоадаптации может привести к новому ароморфозу.
Однако, в большинстве случаев развитие по пути идиоадаптации представляет собой более или менее узкую специализацию и завершается для данной группы «эволюционным тупиком».
Напротив, в ходе ароморфозов идёт накопление не узких адаптаций, а адаптаций наиболее общего универсального значения, которые оказываются полезными во всевозможных условиях (так позвоночник оправдал себя и в воде и на земле и в воздухе).
Дегенерация связана со вторичным упрощением живых существ, (например у паразитических червей) что, впрочем, отнюдь не исключает широкого их распространения и многочисленности.
Северцов настаивал, что все эти направления эволюции являются биологически равноценными, а эволюция - это адаптациогенез и отбор действует сугубо «оппортунистически», приспосабливая организмы к условиям, имеющимся в данное время в данном месте.
При возникновении все адаптации равноценны и лишь впоследствии выясняется, что некоторые из них имеют универсальный ароморфный характер.
В более поздних работах мы находим продолжение этой логики Северцова. Впрочем, ароморфоз уже считается главным направлением эволюционного процесса.
Так, Джулиан Хакали (предложивший сам термин СТЭ) выдвинул концепцию «неограниченного прогресса» (unlimited progress); последний представляет собой биологическое развитие, не препятствующее дальнейшему прогрессу имеющее место в эволюционной линии, ведущей на человека.
Все остальные линии рассматриваются Хакали как «ограниченные» (blind allegs).
Близкую по содержанию концепцию «мегаарогенеза» (или «магистральной линии развития живого») выдвинул К.М. Завадский.
Мегаарогенез- это гигантская цепь ароморфных преобразований, тянущаяся от первичных форм жизни до высших растений, позвоночных и человека; при этом, магистральная линия биологического прогресса рассматривается в качестве результата всей живой материи.
Биологическая эволюция отлична от эволюции атомов, Земли, общества и др. В ее основе — «уникальные процессы самовоспроизведения макромолекул и живых организмов, которые таят в себе почти неограниченные возможности преобразования живых систем в ряду поколений», — отмечает известный эволюционист А.В. Яблоков. Биологическая эволюция — необратимое и в известной степени направленное историческое развитие живой природы, сопровождающееся изменением генетического состава популяций, формированием адаптаций, образованием и вымиранием видов, преобразованиями биогеоценозов и биосферы в целом, отмечает он. С возникновения жизни органическая природа непрерывно развивается сотни миллионов лет, и результатом процесса эволюции является то разнообразие форм живой материи, которая еще не полностью описана, классифицирована и изучена. Формы живой материи — и предмет, и объект эволюции. Результаты биологической эволюции многообразны, но всегда представляют собой определенное соответствие развивающейся живой системы условиям ее существования (Дубнищева Т.Я. Концепции современного естествознания. М.: Академия, 2006. С. 465 – 501).
Основные узлы и механизмы станка.
Станина - является базовой деталью станка и служит для монтажа всех основных частей станка. Должна быть жесткой, виброустойчивой, иметь малую металлоемкость и стоимость. На станине должны быть предусмотрены проемы и окна для сборки и смазки. Она имеет коробчатую форму с ребрами жесткости.
Станины подразделяются на горизонтальные и вертикальные. Станина с вертикальной компоновкой называется стойкой.
В серийном производстве станины изготавливают литыми из серого чугуна.
В единичном производстве станины изготавливают сварными из прокатной стали. Иногда для изготовления станин тяжелых станков применяют железобетон.
Направляющие - наиболее ответственная часть станины. Служащая для перемещения инструмента и заготовки.
В станках применяют направляющие скольжения и качения для прямолинейного и кругового перемещения.
По форме направляющие делятся на:
а) охватываемые - применяются при малых скоростях , на них плохо удерживается смазка, но не пристает стружка.

б) охватывающие - применяются для высоких скоростей скольжения, т. к. хорошо удерживают смазку. Их нужно защищать от стружки и грязи.
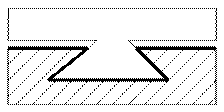
в) комбинированные - это когда одна из направляющих выполнена плоской, а вторая призматической, V - образной или в виде ласточкиного хвоста.
г) накладные - выполненные в виде планок, прикрепляемых винтами к литой чугунной станине или привариваемых к стальной сварной станине.
д)гидростатические - предусматривают подвод масла к сопрягаемым поверхностям под давлением и обеспечивают создание масляной подушки по всей площади контакта
е) аэростатические - предусматривают создание воздушной подушки в зазоре между сопряженными поверхностями направляющих.
В станках также применяют направляющие качения, которые могут быть открытыми и закрытыми.
Дата добавления: 2015-03-20; просмотров: 777;