Строение митохондрий
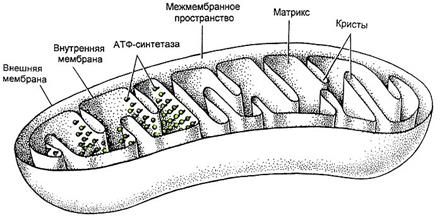 Каждая митохондрия состоит из наружной и внутренней мембран, между которыми находится межмембранное пространство (Рис.7). Внутренняя мембрана образует складки - кристы, обращенные внутрь митохондрии. Пространство, ограниченное внутренней мембраной, заполнено митохондриальным матриксом, - мелкозернистым материалом различной электронной плотности.
Каждая митохондрия состоит из наружной и внутренней мембран, между которыми находится межмембранное пространство (Рис.7). Внутренняя мембрана образует складки - кристы, обращенные внутрь митохондрии. Пространство, ограниченное внутренней мембраной, заполнено митохондриальным матриксом, - мелкозернистым материалом различной электронной плотности.
Рис.7.
Наружная мембрана митохондрий содержит много молекул специализированных транспортных белков (например, порин), что обеспечивает её высокую проницаемость, а также белки-рецепторы, распознающие белки, которые переносятся через обе мембраны митохондрий в особых точках их контакта – зонах слипания.
Внутренняя мембранамитохондрий образует складки – кристы, благодаря чему значительно увеличивается внутренняя поверхность митохондрий. В состав внутренней мембраны входят транспортные белки; ферменты дыхательной цепи и сукцинатдегидрогеназа; комплекс АТФ-синтетазы. На кристах имеются элементарные частицы (оксисомы, или F1-частицы), состоящие из округлой головки (9 нм) и цилиндрической ножки. Именно на них происходит сопряжение процессов окисления и фосфорилирования (АДФ → АТФ).
Чаще всего кристы располагаются перпендикулярно длинной оси митохондрий и имеют пластинчатую (ламеллярную) форму. В клетках,синтезирующих стероидные гормоны, кристы имеют вид трубочек или пузырьков - тубулярно-везикулярные кристы. В этих клетках ферменты стероидного синтеза частично локализуются на внутренней мембране митохондрий
Число и площадь крист отражает функциональную активность клеток: наибольшая площадь крист характерна, например, для митохондрий клеток сердечной мышцы, где потребность в энергии постоянно очень велика.
Митохондриальный матрикс – мелкозернистое вещество, заполняющее полость митохондрии. Матрикс содержит несколько сотен ферментов: ферменты цикла Кребса, окисления жирных кислот, белкового синтеза. Здесь иногда встречаются митохондриальные гранулы, а также локализуются митохондриальные ДНК, иРНК, тРНК, рРНКи митохондриальные рибосомы.Митохондриальные гранулы – частицы высокой электронной плотности диаметром 20-50 нм, содержащие ионы Са и Мg.
Митохондриальная ДНК имеет кольцевую форму и включает 37 генов. Генетическая информация митохондриальной ДНК обеспечивает синтез около 5-6% белков митохондрий (ферменты электрон-транспортной системы). Синтез других митохондриальных белков контролируется ДНК ядра. Наследование митохондриальной ДНК происходит только по материнской линии.
Повреждения митохондриальной ДНК в результате мутаций могут привести к развитию ряда патологий - митохондриальных цитопатий (синдромы Барта, Патерсона, МERRF (красных разорванных волокон) и др.).
ЛИЗОСОМЫ – мембранные органеллы, которые обеспечивают внутриклеточное переваривание (расщепление) макромолекул внеклеточного и внутриклеточного происхождения, и обновление компонентов клетки.
Морфологически лизосомы представляют собой округлые пузырьки, ограниченные мембраной и содержащие большое количество различных гидролаз (более 60 ферментов). Наиболее характерными ферментами лизосом являются: кислая фосфатаза (маркёр лизосом), протеазы, нуклеазы, сульфатазы, липазы, гликозидазы. Все литические ферменты лизосом представляют собой кислые гидролазы, т.е. оптимум их активности проявляется при рН≈5.
Мембрана лизосом (около 6 нм толщиной) обладает протонным насосом, вызывающим закисление среды внутри органелл, обеспечивает диффузию низкомолекулярных продуктов переваривания макромолекул в гиалоплазму и препятствует утечке литических ферментов в гиалоплазму.
Повреждение мембраны приводит к разрушению клетки вследствие самопереваривания.
Лизосомы присутствуют во всех клетках. Особенно много лизосом в тех клетках, где активно протекают процессы фагоцитоза с последующим перевариванием захваченного материала (например, в нейтрофильных гранулоцитах, макрофагах, остеокластах).
Лизосомы подразделяются на первичные (неактивные)ивторичные (активные).
Первичные лизосомы (гидролазные пузырьки) – округлые пузырьки небольшого размера (обычно около 50 нм диаметром), с мелкозернистым, гомогенным, плотным матриксом. Надежная идентификация первичных лизосом возможна только при гистохимическом выявлении характерных ферментов (кислая фосфатаза). Первичные лизосомы – неактивные структуры, еще не вступившие в процессы расщепления субстратов.
Вторичные лизосомы – органеллы, активно участвующие в процессах внутриклеточного переваривания. Диаметр вторичных лизосом обычно составляет 0.5-2 мкм, их форма и структура могут существенно варьировать в зависимости от перевариваемого субстрата, но обычно содержимое вторичных лизосом гетерогенно.
Вторичная лизосома – результат слияния первичной лизосомы с фагосомой или аутофагосомой (Рис.8).
Фаголизосома формируется путем слияния первичной лизосомы с фагосомой - мембранным пузырьком, содержащим материал, захваченный клеткой извне. Процесс разрушения этого материала называется гетерофагией. Гетерофагия играет важную роль в функции всех клеток. Особое значение гетерофагия имеет для клеток, осуществляющих защитную функцию, таких как макрофаги и нейтрофильные лейкоциты, которые захватывают и переваривают болезнетворные микроорганизмы.
Аутофаголизосома образуется при слиянии первичной лизосомы с аутофагосомой - мембранным пузырьком, содержащим собственные компоненты клетки, которые подлежат разрушению. Процесс переваривания внутриклеточного материала называется аутофагией. Аутофагия обеспечивает постоянное обновление клеточных структур благодаря перевариванию митохондрий, полисом, фрагментов мембран.
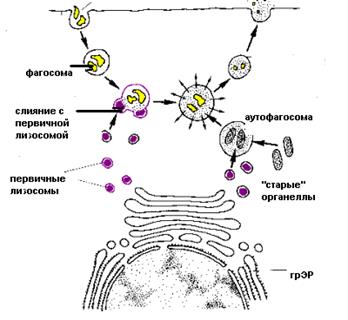
Рис.8.
Остаточные тельца – лизосомы, содержащие непереваренный материал, которые могут находиться в цитоплазме длительное время. В некоторых долгоживущих клетках (нейроны, кардиомиоциты, гепатоциты) в остаточных тельцах накапливается коричневый эндогенный пигмент липофусцин – «пигмент старения».
Дефицит лизосомальных ферментов может приводить к развитию ряда заболеваний (болезни накопления), вызванных накоплением в клетках непереваренных веществ, которые нарушают функцию клеток. Примерами могут служить: болезнь Хюрлера, при которой из-за отсутствия α-L-идуронидазы фибробласты и остеобласты накапливают дерматан сульфат, а у больных отмечаются множественные дефекты хондро- и остеогенеза и умственное отставание; болезнь Тэя-Сакса (из-за недостаточности гексозаминидазы А происходит накопление гликолипидов в нервных клетках и поражается нервная система); болезнь Гоше (вследствие наследственного дефекта глюкоцереброзидазы гликолипиды накапливаются в макрофагах и поражаются печень и селезенка) и другие.
Пероксисомы– сферические мембранные органеллы диаметром 0.05 – 1.5 мкм, с умеренно плотным гомогенным или мелкозернистым матриксом. Мелкие пероксисомы встречаются во всех клетках, а крупные пероксисомы – в гепатоцитах, макрофагах, в клетках канальцев почки. Матрикс пероксисом содержит до 50 различных ферментов, важнейшие из которых: каталаза (маркёр пероксисом), пероксидаза, оксидазы аминокислот, уратоксидаза.
У некоторых видов животных в пероксисомах выявляется более плотная кристаллическая сердцевина – нуклеоид, состоящая из уратоксидазы. В пероксисомах клеток человека нуклеотида нет, поскольку отсутствует способность метаболизировать ураты.
Функции пероксисом:
• окисление аминокислот и других субстратов;
• защита клетки от действия перекиси водорода, сильного окислителя, образующегося в результате окисления органических соединений, и оказывающего повреждающий эффект на клетку. При этом каталаза пероксисом разлагает перекись водорода на воду и кислород.
• участие в расщеплении жирных кислот;
• участие в обезвреживании ряда веществ (спирт и др.).
Нарушения активности пероксисом вызывает ряд наследственных заболеваний – пероксисомных болезней с тяжелыми нарушениями нервной системы (синдром Целльвегера и др.)
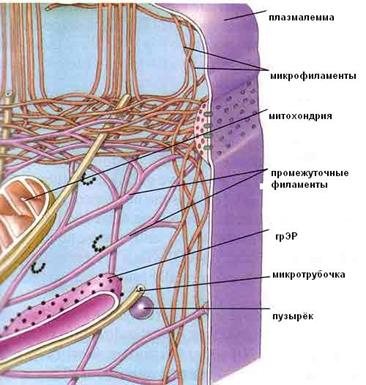
Рис.9.
Цитоскелет – сложная трехмерная сеть немембранных органелл (рис.9):
· микротрубочек;
· микрофиламентов;
· промежуточных филаментов.
Основная функция цитоскелета – опорно-двигательная:
• поддержание и изменение формы клеток;
• перемещение компонентов внутри клетки;
• транспорт веществ внутрь клетки и из клетки;
• обеспечение подвижности клетки
Микротрубочки – наиболее крупные компоненты цитоскелета. Микротрубочки – полые цилиндрические образования различной длины, с диаметром 24-25 нм, с толщиной стенки 5 нм.
 Стенка микротрубочки состоит из спирально
Стенка микротрубочки состоит из спирально
расположенных нитей – профиламентов, образованных димерами из глобулярных белковых молекул – α- и β-тубулина.
Стенка микротрубочки образована 13 субъединицами-профиламентами.
Микротрубочки могут располагаться в цитоплазме в виде отдельных элементов, в виде пучков, где они связаны тонкими поперечными мостиками, или могут частично сливаться друг с другом, образуя дуплеты (в аксонеме ресничек и жгутиков) и триплеты (в базальном тельце и центриолях.
Микротрубочки представляют собой лабильную систему, в которой сохраняется равновесие между их постоянной сборкой и диссоциацией.
Центрами организации микротрубочек (ЦОМТ) являются сателлиты – глобулярные белковые структуры, содержащиеся в базальных тельцах ресничек и клеточном центре, а также центромеры хромосом.
Функции микротрубочек:
· поддержание стабильной формы клеток, и порядка распределения её компонентов;
· обеспечение внутриклеточного транспорта, в том числе органелл, пузырьков, секреторных гранул (благодаря некоторым белкам, ассоциированным с микротрубочками);
· образование основы центриолей и ахроматинового веретена деления и обеспечение движения хромосом в процессе митоза;
· образование основы ресничек и жгутиков, а также обеспечение их движения.
Угнетение самосборки микротрубочек при действии на клетку блокаторов (колхицин и др.) вызывает гибель быстроделящихся клеток вследствие отсутствия митотического веретена деления, нарушения транспортных процессов в клетке (аксонный транспорт в нейронах, секреция), изменения форм клетки, дезорганизацию клеточных органелл (в частности, цистерн ЭПС).
Клеточный центр образован двумя полыми цилиндрическими структурами - центриолями, которые расположены под прямым углом друг к другу.
Каждая центриоль представляет собой короткий цилиндр длиной ~ 0,5 мкм и диаметром ~ 0,2 мкм, состоящий из 9 триплетов частично слившихся трубочек (А, В и С), связанных поперечными белковыми мостиками (рис.10).
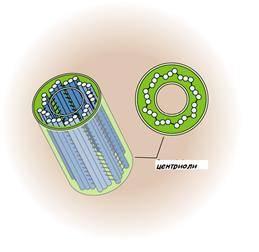
Формула строения центриоли описывается как (9 × 3) + 0, так как в центральной части микротрубочки отсутствуют. Каждый триплет центриоли связан с глобулярными белковыми тельцами – сателлитами, от которых отходят микротрубочки, образующие центросферу.
Рис.10.
В неделящейся клетке выявляется одна пара центриолей – диплосома, которая располагается обычно вблизи ядра. Перед делением клетки в S-периоде интерфазы происходит дупликация центриолей: под прямым углом к каждой зрелой (материнской) центриоли пары образуется новая (дочерняя) центриоль.
В ранней профазе митоза пары центриолей расходятся к полюсам клетки и служат центрами образования микротрубочек ахроматинового веретена деления.
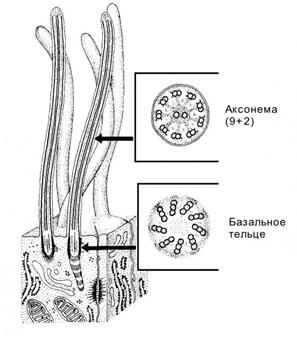
Реснички и жгутикиявляются выростами цитоплазмы, обладающие подвижностью. Основу ресничек и жгутиков составляет каркас из микротрубочек, называемый аксонемой (рис.11).
Длина ресничек равна 2-10 мкм, а их количество на поверхности одной клетки может составлять до нескольких сотен.
В организме человека жгутик есть только в одном типе клеток – сперматозоидах. При этом один сперматозоид имеет один жгутик длиной 50-70 мкм.
Рис.11.
Аксонемаобразована 9периферическими парами микротрубочек (микротрубочки А и В) и одной центрально расположенной парой; такое строение описывается формулой (9 × 2) + 2. Центральная пара микротрубочек окружена центральной оболочкой, от которой к периферическим дуплетам расходятся радиальные спицы. Периферические дублеты связаны друг с другом мостиками белка нексина, а от микротрубочки А к микротрубочке В соседнего дублета отходят “ручки” из белка динеина, который обладает АТФ-азной активностью, что необходимо для скольжения соседних дублетов в аксонеме, вызывающих движение (биение) ресничек и жгутиков
Мутации, вызывающих изменения белков ресничек и жгутиков, ведут к различным нарушениям функций клеток. Так, при отсутствии динеиновых ручек (синдром неподвижных ресничек, или синдром Картагенера), больные страдают хроническими заболеваниями дыхательной системы и бесплодием (вследствие неподвижности спермиев и нарушений продвижения яйцеклеток по яйцеводу).
В основании каждой реснички или жгутика лежит базальное тельце, сходное по строению с центриолью. На уровне апикального конца базального тельца микротрубочка С триплета заканчивается, тогда как микротрубочки А и В продолжаются в соответствующие микротрубочки аксонемы реснички. При развитии ресничек или жгутика базальное тельце играет роль матрицы, на которой происходит сборка компонентов аксонемы
Микрофиламенты – тонкие белковые нити диаметром 5-7 нм, расположенные в цитоплазме поодиночке, в виде сетей или упорядоченными пучками (в скелетной и сердечной мышцах). Основной белок микрофиламентов – актин – встречается в клетках как в мономерной форме (глобулярный G-актин), так и в виде полимерного фибриллярного F-актина.
Функции микрофиламентов:
• в мышечных волокнах и клетках актиновые микрофиламенты образуют упорядоченные пучки и при взаимодействии с миозиновыми филаментами обеспечивают их сокращение.
• в немышечных клетках микрофиламенты образуют кортикальную (терминальную) сеть, в которой микрофиламенты сшиты с помощью особых белков (филамин и др.). Кортикальная сеть, с одной стороны, обеспечивает поддержание формы клетки, а с другой - способствует изменениям формы плазмолеммы, обеспечивая, таким образом, функции эндо- и экзоцитоза, миграции клеток, образования псевдоподий.
• микрофиламенты тесно связаны с органеллами, транспортными пузырьками, секреторными гранулами и играют важную роль в их перемещении внутри цитоплазмы.
• микрофиламенты формируют сократимую перетяжку (срединное тельце) при цитотомии, завершающей клеточное деление.
• микрофиламенты участвуют в организации структуры межклеточных соединений (zonula adherens – поясок сцепления).
• микрофиламенты являются основой специальных выростов цитоплазмы – микроворсинок и стереоцилий.
Микроворсинки– пальцевидные выросты цитоплазмы клетки диаметром 0.1 мкм и длиной 1 мкм, основу которых образуют актиновые микрофиламенты (рис.12).
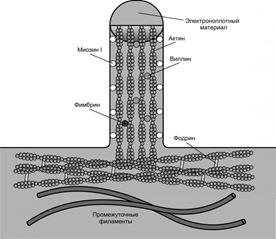 Микроворсинки обеспечивают многократное увеличение площади поверхности клетки. На апикальной поверхности некоторых клеток, активно участвующих в процессах расщепления и всасывания веществ, имеется до несколько тысяч микроворсинок, образующих в совокупности щёточную каемку(эпителий тонкой кишки и почечных канальцев).
Микроворсинки обеспечивают многократное увеличение площади поверхности клетки. На апикальной поверхности некоторых клеток, активно участвующих в процессах расщепления и всасывания веществ, имеется до несколько тысяч микроворсинок, образующих в совокупности щёточную каемку(эпителий тонкой кишки и почечных канальцев).
Рис.12.
Основа каждой микроворсинки – пучок, содержащий около 40 микрофиламентов, расположенных вдоль её длинной оси. Микрофиламенты имеют поперечные сшивки из белков (фимбрин, виллин), и прикреплены к плазмолемме особыми белковыми мостиками (минимиозин). У основания микроворсинки микрофиламенты пучка вплетаются в терминальную сеть
Стереоцилии – длинные, иногда ветвящиеся микроворсинки, имеющие каркас из микрофиламентов. Они встречаются редко (например, в главных клетках эпителия протока придатка семенника).
Промежуточные филаменты – прочные и устойчивые белковые нити толщиной около 10 нм (что является промежуточнымзначением между толщиной микротрубочек и микрофиламентов). Промежуточные филаменты располагаются в виде трехмерных сетей в различных участках цитоплазмы, окружают ядро, участвуют в образовании межклеточных контактов (десмосом) и поддерживают форму отростков.
Дата добавления: 2015-03-19; просмотров: 8779;