ПОПЕРЕЧНОПОЛОСАТАЯ МЫШЕЧНАЯ ТКАНЬ
Из этого вида ткани построены вся соматическая, или скелетная, мускулатура млекопитающих, а также мышцы языка, мышцы, приводящие в движение глазное яблоко, мышцы гортани и некоторые другие. Поперечнополосатые мышцы резко отличаются от гладких тем, что сокращаются они значительно быстрее (доли секунды); это сокращение происходит неритмично; поперечнополосатой ткани свойственна быстрая утомляемость.
Развивается поперечнополосатая мышечная ткань из миотомов, являющихся частью сомитов мезодермы. Миотомы содержат клетки удлиненной формы — миобласты, которые растут, сливаются между собой, образуют многоядерные симпластические образования, называемые мышечными трубочками. Ядра в них располагаются в центре, а в цитоплазме заметна слабая
фибриллярность. В дальнейшем в центральной части мышечных трубочек интенсивно развиваются миофибриллы, а ядра оттесняются к сарколемме. Из окружающей мезенхимы формируется эндомизий, и таким образом окончательно формируется мышечное волокно.
| Поперечнополосатая ткань состоит из поперечнополосатых мышечных волокон, объединенных при помощи рыхлой соединительной ткани в пучки. Мышечные волокна (рис.14) являются неклеточными симпластическими образованиями удлиненной цилиндрической формы. Они имеют длину от нескольких миллиметров до 10—12 см и более. Толщина их колеблется от 10 до 200 мкм и зависит от вида, породы, возраста и физиологической активности животного, а также от типа анатомической структуры мышц. |
|

 В одной мышце наряду с мелкими находятся и крупные волокна. Каждое мышечное волокно покрыто оболочкой — сарколеммой (sarcos — мясо, lemma — оболочка), состоящей из двух основных слоев. Непосредственно к волокну прилегает плазмалемма, аналогичная оболочкам клеток. Наружную часть сарколеммы составляет бесструктурная мембрана, напоминающая базальную мембрану эпителия. Снаружи сарколемму, точнее базальную мембрану, оплетают коллагеновые волокна, которые на некотором расстоянии от мускульного волокна переходят в коллагеновые волокна окружающей соединительной ткани. Содержимое волокна аналогично цитоплазме клеток и называется саркоплазмой.
В одной мышце наряду с мелкими находятся и крупные волокна. Каждое мышечное волокно покрыто оболочкой — сарколеммой (sarcos — мясо, lemma — оболочка), состоящей из двух основных слоев. Непосредственно к волокну прилегает плазмалемма, аналогичная оболочкам клеток. Наружную часть сарколеммы составляет бесструктурная мембрана, напоминающая базальную мембрану эпителия. Снаружи сарколемму, точнее базальную мембрану, оплетают коллагеновые волокна, которые на некотором расстоянии от мускульного волокна переходят в коллагеновые волокна окружающей соединительной ткани. Содержимое волокна аналогично цитоплазме клеток и называется саркоплазмой.
 В саркоплазме находятся ядра, органеллы, а также включения. Ядра в волокне у разных животных располагаются по-разному: у млекопитающих они находятся по периферии волокна под сарколеммой, а у птиц — в центре волокна. В одном волокне может быть свыше сотни ядер. Они имеют форму сильно вытянутых овальных телец и бедны хроматином. В саркоплазме
В саркоплазме находятся ядра, органеллы, а также включения. Ядра в волокне у разных животных располагаются по-разному: у млекопитающих они находятся по периферии волокна под сарколеммой, а у птиц — в центре волокна. В одном волокне может быть свыше сотни ядер. Они имеют форму сильно вытянутых овальных телец и бедны хроматином. В саркоплазме
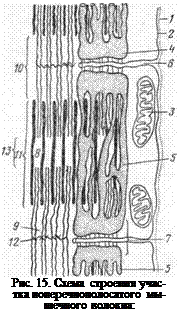
отмечают большое количество крупных митохондрий (саркосом). Особенно много саркосом между миофибриллами. Саркосомы, благодаря содержащимся в них ферментам, принимают активное участие в процессах, связанных с выработкой энергии. Кроме того, в мышечном волокне находится пластинчатый комплекс и аналогичный цитоплазматической сети других клеток саркоплазматический ретикулум — система канальцев, пузырьков, цистерн, располагающихся вдоль волокна, между миофибриллами (рис. 15—4, 5).
Местами сарколемма вдается внутрь волокна, образуя поперечные трубочки — Т-системы или Т-каналы. По ним в волокно поступает вода, и они участвуют в распространении нервного импульса, а также вместе с саркоплазматическим ретикулумом принимают участие в процессе сокращения волокна (6). Комплекс Т-канала и прилегающих к нему с обеих сторон элементов саркоплазматического ретикулума называется триадой.
В саркоплазме поперечнополосатого мышечного волокна содержатся также трофические включения, такие, как жир, гликоген и миоглобин (белок).
Количество жира различно в разных волокнах. От миоглобина зависит цвет мышцы — отсюда красные и белые мышцы. В темно-красных мышцах его больше. Этот белок легко связывает кислород, при его участии происходит дыхательное фосфорилирование, доставляющее большое количество энергии. В более светлоокрашенных мышцах миоглобина меньше, в них преобладает анаэробный процесс обмена углеводов, благодаря чему освобождается меньшее количество энергии. В свете сказанного становится понятным, почему у животных, живущих в условиях кислородной недостаточности, примером которых могут быть водные млекопитающие и обитатели высокогорий, миоглобина особенно много. У диких животных мышцы содержат больше миоглобина, чем у домашних. Мышцы у интенсивно работающего вола окрашены сильнее, чем у менее интенсивно работающего; у молодых животных слабее, чем у взрослых. У кур, потерявших способность летать, грудные мышцы, связанные с движением крыла, окрашены слабо, тогда как активно работающие мышцы тазовых конечностей имеют темно-красный цвет.
 Сократимыми элементами мышечного волокна являются миофибриллы. Каждая миофибрилла представляет собой нить толщиной от 0,5 до 2мкм, а длина соответствует длине волокна. Она состоит из участков, различно преломляющих свет и потому имеющих на препарате вид темных (анизотропных) дисков А и светлых (изотропных) дисков И. В одном волокне миофибриллы располагаются так, что их темные диски приходятся против темных, а светлые против светлых. Через середину каждого изотропного диска проходит полоска Z или полоска Т (телофрагма) (12), а через середину анизотропного — полоска М (мезофрагма). В расслабленной мышце в середине анизотропного диска обнаруживают светлую зону (полоска Н), в центре которой и расположена полоска М. Участок миофибриллы между двумя полосками Z называют саркомером. В него входят половина изотропного диска, целый анизотропный и половина другого изотропного диска. Ввиду того, что миофибрилл в волокне очень много и лежат они очень тесно, под микроскопом различить отдельные фибриллы не удается, и для глаза светлые диски всех миофибрилл сливаются в сплошную поперечную светлую
Сократимыми элементами мышечного волокна являются миофибриллы. Каждая миофибрилла представляет собой нить толщиной от 0,5 до 2мкм, а длина соответствует длине волокна. Она состоит из участков, различно преломляющих свет и потому имеющих на препарате вид темных (анизотропных) дисков А и светлых (изотропных) дисков И. В одном волокне миофибриллы располагаются так, что их темные диски приходятся против темных, а светлые против светлых. Через середину каждого изотропного диска проходит полоска Z или полоска Т (телофрагма) (12), а через середину анизотропного — полоска М (мезофрагма). В расслабленной мышце в середине анизотропного диска обнаруживают светлую зону (полоска Н), в центре которой и расположена полоска М. Участок миофибриллы между двумя полосками Z называют саркомером. В него входят половина изотропного диска, целый анизотропный и половина другого изотропного диска. Ввиду того, что миофибрилл в волокне очень много и лежат они очень тесно, под микроскопом различить отдельные фибриллы не удается, и для глаза светлые диски всех миофибрилл сливаются в сплошную поперечную светлую
полосу, а темные диски — в темную поперечную полосу на мышечном волокне. Отсюда последнее и получило название поперечнополосатого. Под электронным микроскопом обнаружено, что миофибриллы представляют собой пучок протофибрилл (миофиламентов) двух типов (8, 9). Одни из них, более тонкие, берут начало от телофрагмы и состоят из белка актина,
образуют они диски И, но немного заходят и в диски А. Другие, протофибриллы, образуя «зоны перекрывания», более толстые, состоят из миозина и расположены только в диске А. В зонах перекрывания между толстыми (миозиновыми) и тонкими (актиновыми) протофибриллами находятся короткие поперечно ориентированные отростки (мостики). При сокращении тонкие протофибриллы внедряются между толстыми, продвигаясь к мезофрагмам внутрь полоски Н, при этом толстые миозиновые тяжи приближаются к полоскам Z, упираясь в них в конце сокращения, так что диск И как бы исчезает.
При взаимном скольжении актиновых и миозиновых протофибрилл число поперечных мостиков увеличивается, и они активно перемещаются. Величина саркомера при этом уменьшается.
 Количество миофибрилл и саркоплазмы в разных мышечных волокнах различно. В мышцах большинства животных миофибриллы располагаются плотным пучком в середине волокна (плотный тип строения волокна), а у других животных несколькими пучками, разделенными прослойками саркоплазмы (рыхлый тип строения волокна) (рис.16). Группы поперечнополосатых мышечных волокон при помощи соединительной ткани (эндомизий) связываются в пучки I порядка (первичный мышечный пучок). Несколько пучков I порядка более значительным количеством соединительной ткани (внутренний перимизий) объединяются в пучки
Количество миофибрилл и саркоплазмы в разных мышечных волокнах различно. В мышцах большинства животных миофибриллы располагаются плотным пучком в середине волокна (плотный тип строения волокна), а у других животных несколькими пучками, разделенными прослойками саркоплазмы (рыхлый тип строения волокна) (рис.16). Группы поперечнополосатых мышечных волокон при помощи соединительной ткани (эндомизий) связываются в пучки I порядка (первичный мышечный пучок). Несколько пучков I порядка более значительным количеством соединительной ткани (внутренний перимизий) объединяются в пучки
II порядка (вторичный мышечный пучок). Пучки II порядка, соединяясь между собой при помощи новых слоев соединительной ткани, образуют пучки III порядка (третичный мышечный пучок) и т. д. Наконец, самые наружные слои соединительной ткани окутывают всю мышцу (наружный перимизий). Все прослойки соединительной ткани, проходящие между пучками различных порядков, а также между отдельными волокнами в пучке, связаны и составляют единый соединительнотканный каркас — строму мышцы (рис.17). По прослойкам соединительной ткани проходит большое количество кровеносных и лимфатических сосудов, а также нервов. Иннервируется поперечнополосатая мышечная ткань черепно-мозговыми и спинномозговыми нервами. В составе последних находятся как двигательные волокна, передающие возбуждение от головного мозга в мышцу, так и чувствительные, передающие возбуждение от мышцы в головной мозг. Контролируется работа мышц непосредственно корой головного мозга.
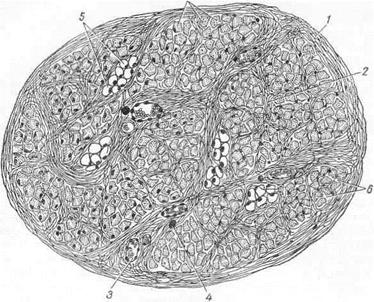 Рис. 17. Схема строения мышцы:
I — наружный перимиэий;_2 — внутренний перимизий; 3 — кровеносные сосуды; 4—нерв; 5 —жировые клетки; 6 — эндомизий; 7 — мышечные волокна в поперечном разрезе (точками обозначены миофибриллы в мышечных волокнах).
Рис. 17. Схема строения мышцы:
I — наружный перимиэий;_2 — внутренний перимизий; 3 — кровеносные сосуды; 4—нерв; 5 —жировые клетки; 6 — эндомизий; 7 — мышечные волокна в поперечном разрезе (точками обозначены миофибриллы в мышечных волокнах).
|
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ
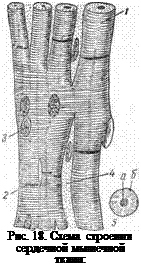 Эта ткань образует один из слоев стенки сердца — миокард. Она делится на собственно сердечную мышечную ткань и проводящую систему.
Эта ткань образует один из слоев стенки сердца — миокард. Она делится на собственно сердечную мышечную ткань и проводящую систему.
Собственно сердечная мышечная ткань по своим физиологическим свойствам занимает промежуточное положение между гладкими мышцами внутренних органов и поперечнополосатыми (скелетными). Она сокращается быстрее гладких, но медленнее поперечнополосатых мышц, работает ритмично и мало утомляется.
 В связи с этим в ее строении имеется ряд своеобразных черт (рис.18). Состоит эта ткань из отдельных мышечных клеток (миоцитов), почти
В связи с этим в ее строении имеется ряд своеобразных черт (рис.18). Состоит эта ткань из отдельных мышечных клеток (миоцитов), почти
прямоугольной формы, расположенных столбиком друг за другом. В целом получается структура, напоминающая поперечнополосатое
волокно, разделенное на отрезки поперечными перегородками — вставочные диски, являющиеся участками плазмалеммы двух соседних
клеток, соприкасающихся друг с другом. Рядом лежащие волокна соединены анастомозами, что позволяет им сокращаться одновременно. Группы мышечных волокон окружены соединительнотканными прослойками, подобными эндомизию. В центре каждой клетки 1—2 ядра овальной формы. Миофибриллы располагаются по периферии клетки и имеют поперечную исчерченность. Между миофибриллами в саркоплазме большое количество митохондрий (саркосом), чрезвычайно богатых кристами, что говорит о высокой их энергетической активности. Снаружи клетка покрыта, кроме плазмалеммы, еще и базальной мембраной. Богатство цитоплазмой и хорошо развитый трофический аппарат обеспечивают сердечной мышце непрерывность деятельности.
Проводящая система сердца состоит из бедных миофибриллами тяжей мышечной ткани способных согласовывать работу разобщенных мышц желудочков и предсердий.
9. НЕРВНАЯ ТКАНЬ
Нервная ткань занимает особое место в организме высокоразвитых животных. Через чувствительные нервные окончания организм получает сведения о внешнем мире. Возбуждение, вызванное такими агентами внешней среды, как звук, свет, температура, химические и прочие воздействия, передается по чувствительным нервным волокнам в определенные участки
центральной нервной системы. Затем нервный импульс в силу определенной, очень сложной организации нервной ткани переходит на другие участки центральной нервной системы. Отсюда он по двигательным волокнам передается к мышцам или железе, которые и осуществляют целесообразную ответную реакцию на раздражение. Она выражается в том, что мышца сокращается, а железа выделяет секрет. Путь от органа чувств до центральной нервной системы и от нее до эффекторного органа (мышца, железа) называется рефлекторной дугой, а сам процесс — рефлексом. Рефлекс — это механизм, при помощи которого животное приспосабливается к меняющимся условиям внешней среды.
На протяжении длительного периода эволюционного развития животных ответная реакция благодаря совершенствованию нервной системы становилась разнообразнее, сложнее, и животные все более и более приспосабливались к различным, часто весьма изменчивым условиям внешней среды.
Особенно сложна и дифференцирована нервная система млекопитающих. У них каждый отдел нервной системы, даже самый небольшой ее участок, имеет свою, только ему свойственную структуру нервной ткани. Однако, несмотря на большое различие нервной ткани разных участков нервной системы, для всех разновидностей ее характерны некоторые общие черты строения. Эта общность заключается в том, что все разновидности нервной ткани построены из нейронов и клеток нейроглии. Нейроны — главная функциональная единица нервной ткани. Именно в них появляется и по ним распространяется нервный импульс. Однако свою деятельность нейрон может осуществлять при тесном контакте с нейроглией. Межклеточного вещества в нервной ткани очень мало и представлено оно межклеточной жидкостью. Глиальные волокна и пластинки относятся к структурным элементам клеток нейроглии, а не к промежуточному веществу ткани.
Нейроглия весьма многофункциональный компонент. Одной из важных функций нейроглии является механическая, так как она образует остов нервной ткани, на котором размещаются нейроны. Другая функция нейроглии — трофическая. Клетки нейроглии играют также защитную роль. Исследования свидетельствуют, что нейроглия косвенно участвует в проведении нервного импульса по нейрону. Нейроглия, по-видимому, обладает также инкреторной функцией.
По происхождению, нейроглию делят на глиоциты и глиальные макрофаги (рис.19).
Глиоциты образуются из того же нервного зачатка, что и нейроны, то есть из нейроэктодермы. Среди глиоцитов различают астроциты, эпиндимоциты и олигодендроглиоциты. Основная клеточная форма из них — астроциты.
В центральной нервной системе опорный аппарат представлен мелкими клетками с многочисленными радиально расходящимися отростками. В специальной литературе различают два вида астроцитов: плазматические и волокнистые. Плазматические астроциты находятся преимущественно в сером веществе головного и спинного мозга. Клетка характеризуется наличием крупного, бедного хроматином ядра. От тела клетки отходят многочисленные короткие отростки. Цитоплазма богата митохондриями, что говорит об участии астроцитов в обменных процессах. Волокнистые астроциты располагаются в основном в белом веществе мозга. Эти клетки имеют до 40 длинных, слабо ветвящихся отростков.
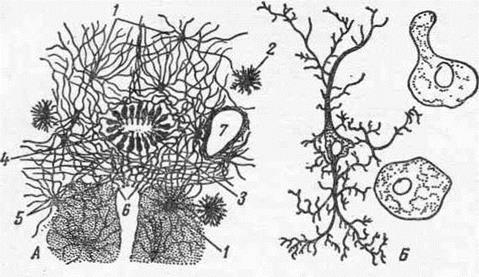 Рис. 19. Глиоциты спинного мозга (А) и глиальные макрофаги (Б):
1 — длиннолучевые, или волокнистые, астроциты; 2 — коротколучевые, или протсплазматнческие, астроциты; 3 — клетки эпендимы; 4 — апикальные концы этих клеток, несущие мерцательные рес- нички, создающие ток цереброспинальной жидкости в желудочках мозга и спинномозговом канале; 5 — отростки клеток эпендимы, образующие остов нервной ткани; 6—концевые пуговки отростков эпендимы, отграничивающие подобно мембране центральную нервную систему от окружающих тканей.
Рис. 19. Глиоциты спинного мозга (А) и глиальные макрофаги (Б):
1 — длиннолучевые, или волокнистые, астроциты; 2 — коротколучевые, или протсплазматнческие, астроциты; 3 — клетки эпендимы; 4 — апикальные концы этих клеток, несущие мерцательные рес- нички, создающие ток цереброспинальной жидкости в желудочках мозга и спинномозговом канале; 5 — отростки клеток эпендимы, образующие остов нервной ткани; 6—концевые пуговки отростков эпендимы, отграничивающие подобно мембране центральную нервную систему от окружающих тканей.
|
Эпиндимоциты выстилают полости желудков и каналов в головном и спинном мозге. Обращенные в просвет полостей и каналов концы клеток несут мерцательные реснички, обеспечивающие ток спинномозговой жидкости. От противоположных концов этих клеток отходят отростки, пронизывающие все вещество мозга. Эти отростки также играют опорную роль.
Олигодендроглиоциты окружают тела невроцитов в центральной и периферической нервных системах, находятся в оболочках нервных волокон. В различных отделах нервной системы они имеют разную форму. От тел этих клеток отходит несколько коротких и слабо разветвленных отростков. Функциональное значение олигодендроглиоцитов очень разнообразно (трофическое, участие в регенерации и дегенерации волокон и т. д.).
Глиальные макрофаги развиваются из клеток мезенхимы, которые при развитии нервной системы проникают в нее вместе с кровеносными сосудами. Глиальные макрофаги состоят из клеток довольно разнообразной формы, но для большинства этих клеток характерно наличие сильно разветвленных отростков. Однако встречаются клетки и округлой формы. Глиальные макрофаги играют трофическую роль и выполняют защитную фагоцитарную функцию.
Нейроны — это высокоспециализированные клетки, образующие звенья рефлекторной дуги. В нейроне совершаются основные нервные процессы: раздражение, которое возникает в результате воздействия на нервные окончания факторов внешней и внутренней среды;
превращение раздражения в возбуждение и передача нервного импульса. Нейроны разных участков нервной системы имеют разные функцию, строение и размер.
По функции различают нейроны чувствительные, двигательные и передаточные. Чувствительные (афферентные) нейроны воспринимают раздражение и передают возникший в результате раздражения нервный импульс в спинной или головной мозг. Передаточные (ассоциативные) нейроны переводят возбуждение с чувствительных нейронов на двигательные. Двигательные (эфферентные) нейроны передают импульс от головного или спинного мозга к мускулатуре, железам и др.
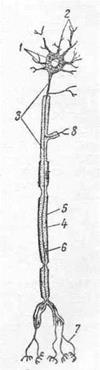 Нейрон состоит из сравнительно компактного и массивного тела и отходящих от него тонких более или менее длинных отростков (рис.20). Тело нервной клетки главным образом управляет ростом и обменными процессами, а отростки осуществляют передачу нервного импульса и вместе с телом клетки ответственны за происхождение импульса. Тело нервной клетки состоит главным образом из цитоплазмы. Ядро бедно хроматином и всегда содержит одно или два хорошо выраженных ядрышка. Из органелл в нервных клетках хорошо развит пластинчатый комплекс, имеется большое количество митохондрий с продольными гребнями. Специфичными для нервной клетки являются базофильное вещество ее и нейрофибриллы (рис. 21).
Нейрон состоит из сравнительно компактного и массивного тела и отходящих от него тонких более или менее длинных отростков (рис.20). Тело нервной клетки главным образом управляет ростом и обменными процессами, а отростки осуществляют передачу нервного импульса и вместе с телом клетки ответственны за происхождение импульса. Тело нервной клетки состоит главным образом из цитоплазмы. Ядро бедно хроматином и всегда содержит одно или два хорошо выраженных ядрышка. Из органелл в нервных клетках хорошо развит пластинчатый комплекс, имеется большое количество митохондрий с продольными гребнями. Специфичными для нервной клетки являются базофильное вещество ее и нейрофибриллы (рис. 21).

 Базофильное, или тигроидное, вещество состоит из белковых веществ, содержащих железо и фосфор. Оно богато рибонуклеиновой кислотой и гликогеном. В виде глыбок неправильной формы это вещество разбросано по всему телу клетки и
Базофильное, или тигроидное, вещество состоит из белковых веществ, содержащих железо и фосфор. Оно богато рибонуклеиновой кислотой и гликогеном. В виде глыбок неправильной формы это вещество разбросано по всему телу клетки и
придает ей пятнистый вид (I). В живой неокрашенной клетке этого вещества не видно. Электронная микроскопия показала, что базофильное вещество идентично зернистой цитоплазматической сети и состоит из сложной сети мембран, которые формируют трубочки или цистерны, лежащие параллельно друг другу и связанные в единое целое. На стенках мембран располагаются гранулы — рибосомы (диаметр 100—300 А), богатые РНК. С базофильным веществом связаны важнейшие физиологические процессы, совершающиеся в клетке. Известно, например, что при утомлении нервной системы количество тигроидного вещества резко уменьшается, а во время отдыха оно восстанавливается.
Нейрофибриллы на фиксированных препаратах имеют вид тонких нитей, расположенных в теле клетки довольно беспорядочно (II). Электронный микроскоп показал, что фибриллярные элементы нервной клетки, аксона и дендритов состоят из трубочек диаметром 200—300 А. Обнаруживают также более тонкие нити — нейрофиламенты, толщина 100 А. При изготовлении препаратов они могут объединяться в пучки, видимые в световом микроскопе в виде нейрофибрилл. Функция их, вероятно, связана с трофическими процессами.
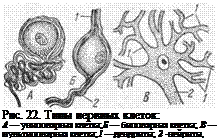 Отростки нервной клетки проводят возбуждение со скоростью около 100 м/с. В зависимости от количества отростков различают нейроны: униполярные — с одним отростком, биполярные — с двумя отростками, ложноуниполярные — развиваются из биполярных, но во взрослом состоянии имеют один отросток, слившийся из двух ранее самостоятельных отростков, и мультиполярные — с несколькими отростками (рис.22). У млекопитающих
Отростки нервной клетки проводят возбуждение со скоростью около 100 м/с. В зависимости от количества отростков различают нейроны: униполярные — с одним отростком, биполярные — с двумя отростками, ложноуниполярные — развиваются из биполярных, но во взрослом состоянии имеют один отросток, слившийся из двух ранее самостоятельных отростков, и мультиполярные — с несколькими отростками (рис.22). У млекопитающих
чувствительные нейроны являются ложноуниполярными (за исключением клеток Догеля II типа), и их тела лежат либо в спинномозговых ганглиях, либо в чувствительных черепно-
мозговых нервах. Передаточные и двигательные нейроны являются мультиполярными. Отростки одной нервной клетки не равнозначны. На основе функции различают два вида отростков: нейрит и дендриты.
Нейритом или аксоном называется отросток, по которому возбуждение передается от тела клетки, то есть центробежно. Он является обязательной ставной частью нервной клетки. От тела каждой клетки отходит только один нейрит, который по длине может варьировать от нескольких миллиметров до 1,5 м, а по толщине от 5 до 500 мкм (у кальмара), но у млекопитающих чаще диаметр колеблется около 0,025 нм (нанометр, миллимикрон).
Разветвляется нейрит обычно сильно лишь на самом конце. На остальном протяжении от него отходят немногочисленные боковые веточки (коллатерали). Благодаря этому диаметр аксона уменьшается незначительно, что обеспечивает большую скорость нервного импульса. В аксоне находятся протонейрофибриллы, но в них никогда не встречается базальное вещество.
Дендриты — отростки, которые в отличие от аксона воспринимают раздражение и передают возбуждение к телу клетки, то есть центростремительно. У очень многих нервных клеток эти отростки древовидно ветвятся, что и дало повод назвать их дендритами (dendron — дерево). В дендритах имеются не только протонейрофибриллы, но и базофильное вещество. От тела мультиполярных клеток отходит несколько дендритов, от тела биполярной — один, а униполярная клетка лишена дендритов. В этом случае раздражение воспринимается телом клетки.
| Рис. 23. Строение безмиелинового нервного волокна: |
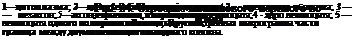 | |||
 |
Нервное волокно — отросток нервной клетки, окруженный оболочками (рис.23,24). Цитоплазматический отросток нервной клетки, занимающий центр волокна, называется осевым цилиндром. Он может быть представлен либо дендритом, либо нейритом. Оболочка нервного волокна построена за счет леммоцита. От толщины осевого цилиндра и строения оболочек волокна зависит скорость передачи нервного импульса, которая колеблется от нескольких м/с до 90, 100 и может достигать 5000 м/с. В зависимости от строения оболочек различают нервные волокна безмиелиновые и миелиновые. И в тех и в других волокнах оболочка, окружающая цитоплазматический отросток нервной клетки, состоит из леммоцитов, но морфологически отличающихся друг от друга. Безмиелиновые волокна представляют собой несколько осевых цилиндров, принадлежащих разным нервным клеткам, погруженных в массу леммоцитов. Эти клетки лежат друг над другом вдоль волокна. Осевые цилиндры могут переходить из одного волокна в другое, а иногда глубоко внедряться в леммоциты, увлекая за собой их плазмалемму.
Благодаря этому образуются мезаксоны (рис.23—4). По безмиелиновым волокнам нервный импульс проходит медленнее и может передаваться лежащим рядом с ними отросткам других нейритов, а благодаря переходу осевых цилиндров из одного волокна в другое передача возбуждения имеет нестрого направленный, а разлитой, диффузный характер. Безмиелиновые волокна находятся главным образом во внутренних органах, которые осуществляют свою функцию сравнительно медленно и диффузно.
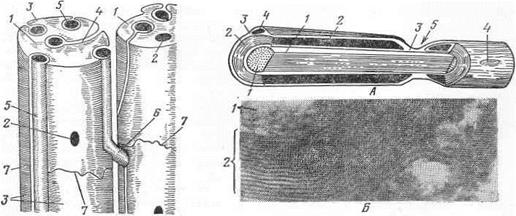
Миелиновые волокна отличаются от безмиелиновых большой толщиной и усложненным строением оболочки (рис.24). В процессе развития отросток нервной клетки, называемый в волокне осевым цилиндром, погружается в леммоцит (шванновскую клетку). В результате вначале он облекается одним слоем плазмалеммы леммоцита, состоящей, как и оболочки других клеток, из бимолекулярного слоя липидов, располагающихся между мономолекулярными слоями белков. Дальнейшее внедрение осевого цилиндра приводит к образованию мезаксона, аналогичного таковому безмиелинового волокна. Однако в случае развития миелинового волокна вследствие удлинения мезаксона и наслоения его вокруг осевого цилиндра (рис.23) развивается многослойная оболочка, называемая миелиновой (рис.25). Благодаря присутствию большого количества липидов она хорошо импрегнируется осмием, после чего ее легко можно увидеть в световой микроскоп. Миелиновая оболочка служит изолятором, благодаря которому нервное возбуждение не может переходить на соседнее волокно. По мере развития миелиновой оболочки цитоплазма леммоцитов оттесняется ею и образует очень тонкий поверхностный слой, называемый неврилеммой. В ней лежат ядра леммоцитов. Таким образом, и миелиновая оболочка и неврилемма являются производными леммоцитов.
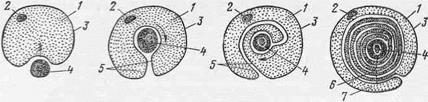 Рис. 25. Схема развития миелинового волокна:
1 — леммоцит; 2 — его ядро; 3 — его плазмалемма; 4 — осевой цилиндр; 5 — мезаксон; стрелкой указано направление вращения осевого цилиндра; 6—будущая миелиновая оболочка нервного волокна;7 — неврилемма, его же.
Рис. 25. Схема развития миелинового волокна:
1 — леммоцит; 2 — его ядро; 3 — его плазмалемма; 4 — осевой цилиндр; 5 — мезаксон; стрелкой указано направление вращения осевого цилиндра; 6—будущая миелиновая оболочка нервного волокна;7 — неврилемма, его же.
|
 Миелиновая оболочка нервных волокон, проходящих в белом веществе спинного и головного мозга, а также в периферических нервах белых мышц у птиц, имеет вид сплошного цилиндра.
Миелиновая оболочка нервных волокон, проходящих в белом веществе спинного и головного мозга, а также в периферических нервах белых мышц у птиц, имеет вид сплошного цилиндра.
В нервных волокнах, составляющих большинство периферических нервов, она прерывается, то есть состоит из отдельных муфт, между которыми имеются промежутки — перехваты Ранвье. В последнем леммоциты соединяются друг с другом. Осевой цилиндр здесь покрыт лишь неврилеммой. Это облегчает поступление питательных веществ в отросток нервной клетки.
Биофизики полагают, что перехваты Ранвье способствуют более ускоренному проведению нервного импульса по отростку, являясь местом регенерации электрического сигнала. Миелиновая оболочка, заключенная между перехватами Ранвье (сегмент), пересекается воронкообразными щелями — миелиновыми насечками, идущими в косом направлении от наружной поверхности оболочки к внутренней. Число насечек в сегменте различное. В миелиновых волокнах возбуждение проводится быстрее и не переходит на соседние волокна.
Нерв. Нервные волокна в головном и спинном мозге составляют главную массу белого
вещества. Выходя из мозга, эти волокна идут не
изолированно, а объединяются друг с другом при
помощи соединительной ткани. Такой комплекс
нервных волокон называют нервом (рис.26). В состав нерва входит от нескольких тысяч до нескольких миллионов волокон. Они
образуют один или несколько пучков — стволиков. В пучки волокна объединяются при помощи соединительной ткани, называемой эндоневрием. Снаружи каждый пучок окружен периневрием. Последний иногда состоит из нескольких слоев плоских эпителиоподобных нейроглиального происхождения клеток и из соединительной ткани, а в других случаях построен только из соединительной ткани. Периневрий играет защитную роль. Несколько таких пучков объединяются друг с другом при помощи более плотной соединительной ткани, называемой эпиневрием. Последний покрывает весь нерв снаружи и служит для укрепления нерва в определенном положении. По соединительной ткани в нерв вступают кровеносные и лимфатические сосуды.
Нервные волокна, составляющие нерв, различны по функции и по строению. Если в нерве имеются отростки только двигательных клеток, — это нерв двигательный, если имеются отростки чувствительных клеток — чувствительный, а если и те и другие — смешанный. Нерв образует и миелиновые, и безмиелиновые волокна. Количество их в разных нервах различно. Так в нервах конечностей больше миелиновых волокон, а в межреберных безмиелиновых.
Синапсы — место соединения отростков двух нервных клеток между собой (рис. 27). Нейроны либо прикасаются друг с другом своими отростками, либо отросток одного нейрона соприкасается с телом клетки другого нейрона. Соприкасающиеся концы нервных отростков могут иметь форму вздутий, петелек или оплетать, подобно лианам, другой нейрон и его отростки. Электронно-микроскопические исследования показали, что в синапсе следует различать: два полюса, синаптическую щель между ними и замыкающее утолщение.
Первый полюс представлен концом аксона первой клетки, причем плазмалемма его образует пресинаптическую мембрану. Около нее в аксоне скапливается много митохондрий, иногда присутствуют кольцеобразно расположенные пучки нитей (нейрофиламенты) и всегда находится большое количество синаптических пузырьков. Последние, по-видимому, содержат химические вещества — медиаторы, выделяющиеся в синаптическую щель, и оказывают действие на второй полюс синапса.
Второй полюс образуется либо телом, либо дендритом, либо шиповидным выростом его, либо даже аксоном второго нейрона. Полагают, что в последнем случае происходит торможение, а не возбуждение второго нейрона. Плазмалемма второй нервной клетки формирует второй полюс синапса — постсинаптическую мембрану, отличающуюся большей толщиной. Предполагают, что в ней совершается разрушение медиатора, который возник во
время одиночного импульса. В местах соприкосновения пре- и постсинаптических мембран на них имеются утолщения, которые, по-видимому, укрепляют синаптическую связь. Описаны синапсы без синаптической щели. В этом случае нервный импульс, вероятно, передается без участия медиаторов.
Через синапсы возбуждение может проходить только в одном направлении. Благодаря синапсам нейроны, соединяясь друг с другом, образуют рефлекторную дугу.

 Нервные окончания являются окончаниями нервных волокон, которые благодаря особой структуре могут либо воспринимать раздражение, либо вызывать сокращение мускула или выделение секрета в железе. Окончания или, вернее, начала чувствительных отростков клеток в органах и тканях, воспринимающих раздражения, называют чувствительными нервными окончаниями или рецепторами. Окончания двигательных отростков нейронов, разветвляющиеся в мышцах или железах, называют двигательными нервными окончаниями или эффекторами. Рецепторы делятся на экстерорецепторы, воспринимающие раздражение из внешней среды, проприорецепторы, несущие возбуждение от органов движения, и интерорецепторы, воспринимающие раздражение от внутренних органов. Рецепторы обладают повышенной чувствительностью к определенным видам раздражений. Соответственно этому имеются механорецепторы,
Нервные окончания являются окончаниями нервных волокон, которые благодаря особой структуре могут либо воспринимать раздражение, либо вызывать сокращение мускула или выделение секрета в железе. Окончания или, вернее, начала чувствительных отростков клеток в органах и тканях, воспринимающих раздражения, называют чувствительными нервными окончаниями или рецепторами. Окончания двигательных отростков нейронов, разветвляющиеся в мышцах или железах, называют двигательными нервными окончаниями или эффекторами. Рецепторы делятся на экстерорецепторы, воспринимающие раздражение из внешней среды, проприорецепторы, несущие возбуждение от органов движения, и интерорецепторы, воспринимающие раздражение от внутренних органов. Рецепторы обладают повышенной чувствительностью к определенным видам раздражений. Соответственно этому имеются механорецепторы,
хеморецепторы и т. д. По строению рецепторы бывают простыми, или свободными, и инкапсулированными.
Свободные нервные окончания (рис.28). Проникнув в ткань, нервное волокно чувствительного нерва освобождается от своих оболочек, и осевой цилиндр, многократно разветвляясь, свободно
оканчивается в ткани отдельными веточками, или эти веточки, переплетаясь, образуют сети и клубочки. В эпителии «пятачка» свиньи чувствительные веточки заканчиваются дискоидальными расширениями, на которых, как на блюдечках, лежат особые чувствительные клетки (меркелевские).
Инкапсулированные нервные окончания очень разнообразны, но в принципе построены одинаково. В таких окончаниях чувствительное волокно освобождается от оболочек, и голый осевой цилиндр распадается на ряд веточек. Они погружаются во внутреннюю колбу, которая состоит из видоизмененных леммоцитов. Внутренняя колба окружена наружной колбой,
состоящей из соединительной ткани.
В поперечнополосатой мышечной ткани чувствительные волокна оплетают сверху мышечные волокна, не проникая внутрь их, и образуют подобие веретена. Сверху веретено покрыто соединительнотканной капсулой.
Двигательные нервные окончания, или эффекторы, в гладкой мышечной ткани и железах обычно построены по типу свободных нервных окончаний. Хорошо изучены моторные окончания в поперечнополосатых мышцах. В месте проникновения двигательного волокна сарколемма мышечного волокна прогибается и одевает голый осевой цилиндр, распадающийся в этом месте на несколько веточек с утолщениями на концах.
|
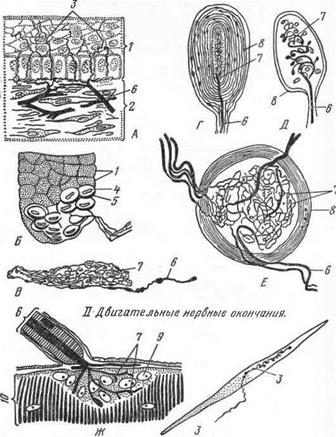 Рис. 28. Типы нервных окончаний:
/ — чувствительные нервные окончания — неинкапсулированные; А — в эпителии роговицы; Б — в эпителии «пятачка» свиньи. В - в перикарде лошади; инкапсулированные. Г — Фатер-Починиево тельце; Д— тельце Майснера; Е—тельце из соска овцы; // —двигательные нервные окончания; Ж - в поперечнополосатом волокне; 3 — в гладкой мышечной клетке; /— эпителий; 2— соединительная ткань; 3— нервныеокончания; 4 — меркедевская клетка; 5 — дискоидальное концевое расширение нервного окончания; 6 — нервное волокно; 7 — разветвление осевого цилиндра; 8 — капсула; 9—ядро леммоцита; 10—мышечное волокно.
Рис. 28. Типы нервных окончаний:
/ — чувствительные нервные окончания — неинкапсулированные; А — в эпителии роговицы; Б — в эпителии «пятачка» свиньи. В - в перикарде лошади; инкапсулированные. Г — Фатер-Починиево тельце; Д— тельце Майснера; Е—тельце из соска овцы; // —двигательные нервные окончания; Ж - в поперечнополосатом волокне; 3 — в гладкой мышечной клетке; /— эпителий; 2— соединительная ткань; 3— нервныеокончания; 4 — меркедевская клетка; 5 — дискоидальное концевое расширение нервного окончания; 6 — нервное волокно; 7 — разветвление осевого цилиндра; 8 — капсула; 9—ядро леммоцита; 10—мышечное волокно.
Используемая литература:
Глаголев П. А. и Ипполитова В. И.
«Анатомия сельскохозяйственных животных с основами гистологии и эмбриологии.» Под ред. И. А. Спирюхова и В. Ф. Вракина. Изд. 4-е, перераб. и доп. М., „Колос”, 1977г.
Дата добавления: 2014-12-17; просмотров: 5437;