Хромосомные мутации
Хромосомные мутации связаны с перестройками хромосом и нарушением их структуры, возникающими при делении клеток или в результате физических воздействий. Открытие хромосомных перестроек дало в руки исследователей новый метод анализа генотипа и изучения локализации генов в хромосоме. Хромосомные перестройки играют важную роль в эволюции генотипа, представляя механизм перекомбинации генов как внутри хромосомы, так и между негомологичными хромосомами; они обеспечивают дополнительную перекомбинацию в генотипе.
Хотя хромосомы способны восстанавливать свою структуру, но иногда такие нарушения сохраняются и дают начало новым клеткам и организмам с хромосомными перестройками, получившими название хромосомных аберраций.
С цитологической точки зрения аберрации подразделяются на хромосомные и хроматидные. Это зависит от того, когда возникают перестройки – до или после репликации хромосом. В зависимости от характера возникающих перестроек различают внутри- и межхромосомные аберрации (рис. 43).
Внутрихромосомные перестройки подразделяют на дефишенси, или концевые нехватки; делеции – выпадение средних частей хромосомы, дупликации, или удвоения (умножения) части хромосомы; инверсии – изменения чередования генов в хромосоме вследствие поворота участка хромосомы на 180°. Межхромосомные перестройки включают транслокации – перемещения части одной хромосомы на другую, не гомологичную ей.
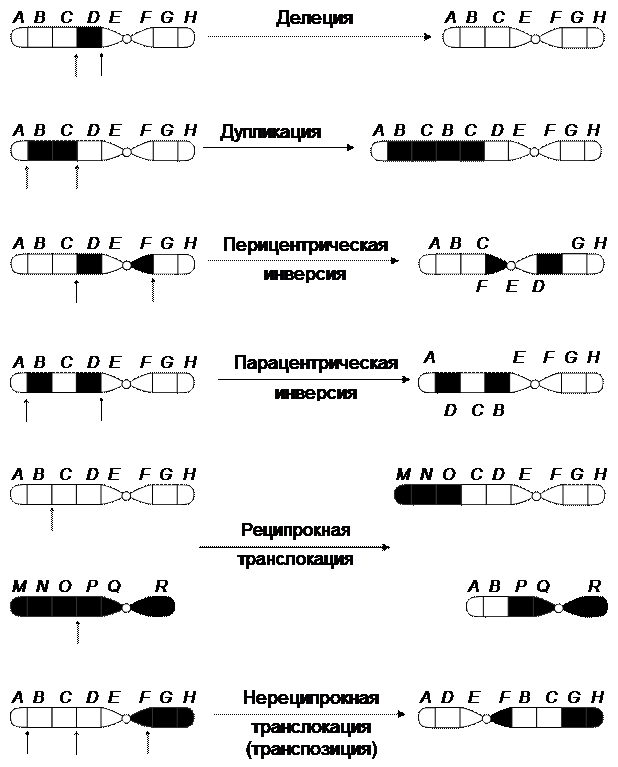 |
Рис. 43. Хромосомные перестройки
Особое положение занимают транспозиции и инсерции – изменения локализации небольших участков генетического материала, включающих один или несколько генов. Транспозиции могут происходить как между негомологичными хромосомами, так и в пределах одной хромосомы. Поэтому транспозиции занимают промежуточное положение между внутри- и межхромосомными перестройками.
Делеции и дефишенси. Эти перестройки хромосом относятся к нехваткам. Вследствие нехваток хромосомы укорачиваются и физическое отсутствие участка одного из гомологов приводит к гемизиготному состоянию генов, находящихся в нормальном гомологе. Если теряются доминантные аллели одного из гомологов гетерозиготы, то наблюдается фенотипическое проявление рецессивных аллелей хромосомы, не затронутой аберрацией. Нехватки, возникающие в результате двух одновременных разрывов в средине хромосомы и элиминации внутреннего участка, называют делециями. Места разрывов соединяются и хромосома становится короче.
Цитологически делеции можно выявить по появлению петли при конъюгации гомологичных хромосом в мейозе. Делеции позволяют определять точное положение изучаемых генов на хромосомных картах.
Если разрыв происходит в одном из плеч хромосомы и концевой участок утрачивается, то такие нехватки относят к терминальным (концевым), или дефишенси. Разрывы иногда происходят одновременно в двух плечах хромосомы, вследствие чего элиминируют оба ее конца. При этом открытые концы могут соединиться, образуя в митозе кольцеобразную хромосому (рис. 44).
 |
Рис. 44. Типы нехватки хромосом:
1 - концевая нехватка;
2 - две концевые нехватки, приводящие к образованию кольца;
3 - внутрихромосомная нехватка.
Нехватки могут быть большими и малыми. К малым относят так называемые микроделеции, которые можно обнаружить только при изучении гигантских хромосом. Микроделеции имитируют генные мутации по фенотипическому эффекту, но отличаются от них только по отсутствию эффекта обратного мутирования. Большие нехватки, как правило, в гомозиготном состоянии летальны, так как нарушают генный баланс. Жизнеспособными могут быть только гетерозиготы по нехваткам. В этом случае нехватки проявляются как доминантные гены.
Большие возможности для выявления делеций, дефишенси и других хромосомных перестроек открывает метод дифференциальной окраски хромосом. Некоторые красители дифференциально окрашивают разные участки хромосом. Благодаря этому хромосомы приобретают характерную поперечную исчерченность. Таким методом определяют хромосомные перестройки на метафазных хромосомах.
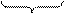
 Дупликации. Эта перестройка связана с двухкратным повторением одного и того же участка хромосомы. Известны случаи многократных повторений, или мультипликаций, какого-либо участка. Их называют также амплификациями. Дупликации могут происходить в пределах одной и той же хромосомы или сопровождаться переносом копии участка генетического материала на другую хромосому (транспозиции). Повторы, возникшие в одной хромосоме, могут располагаться тандемно (АВСВСDЕ…) или инвертированно (АВССВDЕ…). Различают также терминальные повторы, если дупликация затрагивает конец хромосомы.
Дупликации. Эта перестройка связана с двухкратным повторением одного и того же участка хромосомы. Известны случаи многократных повторений, или мультипликаций, какого-либо участка. Их называют также амплификациями. Дупликации могут происходить в пределах одной и той же хромосомы или сопровождаться переносом копии участка генетического материала на другую хромосому (транспозиции). Повторы, возникшие в одной хромосоме, могут располагаться тандемно (АВСВСDЕ…) или инвертированно (АВССВDЕ…). Различают также терминальные повторы, если дупликация затрагивает конец хромосомы.
Многие дупликации и делеции возникают при неравном кроссинговере. Когда в соседних участках хромосомы оказываются похожие последовательности ДНК, то конъюгация гомологов может произойти неправильно. Кроссинговер в таких неправильно конъюгировавших участках хромосом приводит к образованию гамет с дупликацией или делецией.
Цитологически гетерозиготы по дупликациям выявляются сходно с гетерозиготами по делециям – образуется петля при конъюгации хромосом. Дупликации обычно не оказывают такого отрицательного влияния на жизнеспособность, как делеции и дефишенси. Сходные элементы часто повторяются в геномах различных организмов.
Дупликации играют существенную роль в эволюции генома, поскольку они создают дополнительные участки генетического материала, функция которых может быть изменена в результате мутаций и последующего естественного отбора.
Инверсии. Инверсией называют поворот на 180° отдельных участков хромосомы, при этом число генов в хромосоме не изменяется. Если последовательность генов в исходной хромосоме обозначить АВСDЕ, и инверсии подвергся участок ВСD, то в новой хромосоме гены будут расположены в последовательности АDСВЕ.
По отношению к центромере инверсии делят на перицентрические, захватывающие центромеру, и включающие ее в инвертированный участок, и парацентрические, не включающие центромеру в инвертированный участок. В организмах, гомозиготных по хромосомной инверсии, меняется последовательность сцепления генов. У гомозиготы по инверсии АDСВЕ ген А тесно сцеплен с геном D, а не с В, как в исходной хромосоме АВСDЕ, а ген Е сцеплен с В, а не с D.
Инверсии – это широко распространенный путь эволюционного преобразования генетического материала. Например, человек и шимпанзе отличаются по числу хромосом: у человека 2n=46, а у шимпанзе 2n=48. У человека 2-я хромосома содержит большую часть материала, гомологичного дополнительной паре хромосом шимпанзе. Кроме того, различия касаются четырех хромосом: 4-й, 5-й, 12-й и 17-й, в которых произошли перицентрические инверсии. У человека 17-я хромосома акроцентрична, тогда как эта же хромосома у шимпанзе – метацентрик.
У гетерозигот по инверсиям на цитологических препаратах обнаруживают характерные петли – результат конъюгации измененной и нормальной хромосомы. При этом гены очень близко пригоняются друг к другу. Следовательно, между гомологичными локусами хромосом существует взаимное притяжение огромной силы.
Инверсии имеют эволюционное значение. Некоторые близкие виды характеризуются наличием инверсий в хромосомах. Так, в природе было установлено существование двух рас одного вида дрозофилы, не скрещивающихся между собой, причем причина бесплодия заключалась в специфической инверсии у каждой из них. Нескрещиваемость таких рас – это, по существу, начало дивергенции вида, так как мутации, возникающие у одной расы, не могут быть переданы другой; следовательно, их эволюция должна идти разными путями.
Транслокации. Транслокация представляет собой реципрокный обмен участками негомологичных хромосом. Они относятся к межхромосомным перестройкам, которые изменяют группы сцепления генов. Если изобразить последовательность генов в исходных хромосомах, как АВСDЕF и KLMNO, то в транслоцированных хромосомах последовательностями генов могут быть, например, ABCDNO и KLMEF. У гомозигот по этим транслокациям, по сравнению с исходными хромосомами, изменяется характер сцепления: гены, в исходных хромосомах не сцепленные, оказываются сцепленными, и наоборот. Так, гены NO оказываются сцепленными с ABCD и перестают быть сцепленными с KLM. В гетерозиготе по транслокации гены, принадлежащие к разным, негомологичным хромосомам, наследуются как принадлежащие к одной группе сцепления. Это объясняется тем, что полностью функциональными оказываются только те споры (гаметы), которые несут родительские сочетания хромосом.
Характер конъюгации транслоцированных хромосом меняется: образуется фигура креста. Плотная конъюгация вблизи точек разрывов оказывается затрудненной, что приводит к подавлению кроссинговера в этих участках.
На цитологических препаратах у гетерозигот в профазе мейоза образуются квадриваленты, а не биваленты, как обычно, так как гомологичные участки оказываются у всех четырех хромосом. Когда хиазмы сползают от центромер к концам хромосом, крест трансформируется в кольцо. Иногда хромосомы кольца переворачиваются и образуют фигуры типа восьмерки. Из шести возможных типов гаплоидных гамет только два функционируют нормально – те, которые получают полные наборы генов, характерные для исходных родительских форм. Остальные четыре типа гамет несут дупликации и делеции и потому, как правило, не дают жизнеспособного потомства или не участвуют в оплодотворении.
Гетерозиготы по танслокациям частично стерильны, поскольку в процессе мейоза продуцируют дефектные гаметы. Подобно инверсиям, транслокации обеспечивают изоляцию форм и способствуют дивергенции в пределах вида.
В некоторых случаях возможно и практическое использование транслокаций, например, у тутового шелкопряда. Известно, что выход шелковых нитей выше у коконов мужского пола. Поэтому в шелководстве выгоднее выращивать гусениц мужского пола. Для решения этой задачи российскому генетику В.А. Струнникову удалось экспериментальным путем создать мутантную линию, в которой все самцы постоянно гетерозиготны по двум неаллельным рецессивным летальным генам (l1 и l2), локализованным в разных Z-хромосомах самца. Эти самцы вполне жизнеспособны, так как проявление обоих летальных генов подавлено их доминантными аллелями (L1 и L2). При скрещивании самцов с транслоцированными в половые хромосомы летальными генами с нормальными самками в их потомстве выживают только мужские особи, а женские зиготы (WZ) погибают еще в яйце, так как единственная Z-хромосома этих зигот происходит от отца и несет либо один летальный ген (l1), либо другой (l2), а в хромосоме W отсутствуют доминантные аллели этих генов. Получение гусениц мужского пола дает высокий экономический эффект.
Транспозиции и инсерции. Транспозиции представляют собой перемещение небольших участков генетического материала в пределах одной хромосомы или между разными хромосомами. Транспозиции происходят при участии особых подвижных или мигрирующих генетических элементов. Они бывают двух типов: инсерции – относительно короткие последовательности ДНК, которые несут информацию, необходимую для собственной транспозиции, и транспозоны, которые кроме информации, необходимой для транспозиции, кодируют фенотипические признаки.
Первый подвижный (мобильный, транспозирующийся) генетический элемент был описан в 1947 г. Б. Мак-Клинток на кукурузе. Ею был обнаружен мигрирующий локус Ds (диссоциатор), в котором часто происходят разрывы хромосом.
Сам по себе локус Ds не вызывает разрывов. Они появляются в нем, если только в геноме присутствует другой мигрирующий элемент – Ac (активатор). Оба эти элемента могут теряться с частотой нескольких процентов в мейотическом потомстве или менять свою локализацию при митотических делениях. При этом Ds перемещается только в присутствии Ас.
Внедрение Ds в непосредственной близости или внутри гена С, контролирующего красную окраску алейрона семян, приводило к инактивации этого гена, и гетерозиготные семена оказывались неокрашенными. В присутствии Ас диссоциатор Ds начинал перемещаться – иногда покидал локус С. В результате этого появлялись окрашенные пятна алейрона на неокрашенных семенах.
Контролирующие элементы представляют собой класс мутаций, вызывающих обесцвечивание зерен. Эти мутации были названы контролирующими элементами, так как они контролируют активность гена, ответственного за синтез пигмента в данной клетке зерна. Они иногда ревертируют к дикому типу и бывает, что реверсия к дикому типу одного мутантного гена сопровождается возникновением такой же мутации в другом гене. Можно было предположить, что контролирующий элемент «перепрыгивает» из одного антоцианового локуса в другой, то есть Мак-Клинток описала способность контролирующих элементов перемещаться по геному.
Только в 80-х годах ХХ века благодаря успехам генной инженерии удалось выделить и исследовать Ас, Ds и другие мигрирующие элементы у кукурузы. Оказалось, что Ds - это дефектный делетированный вариант Ас. Структура элемента Ас оказалась типичной для мигрирующих элементов, которые были изучены у бактерий, дрозофилы и дрожжей.
Начало изучению молекулярной структуры мигрирующих генетических элементов положило открытие множества необычных мутантов у кишечной палочки (E. coli). Общими для всех мутантов были инсерции большей или меньшей длины. Эти встраивающиеся в разные участки генома E. coli молекулы ДНК получили наименование IS-элементов (от английского insertion sequences – инсерционные последовательности). Было обнаружено, что несколько различных встраивающихся последовательностей могут вызывать мутации многих генов. Они различаются размером, но имеют некоторые общие черты строения. На концах содержатся одинаковые или почти одинаковые нуклеотидные последовательности, расположенные, однако, в обратном порядке. Кроме того, когда инсерция встраивается в ДНК-мишень, небольшой участок последовательности ДНК-мишени повторяется около каждого конца инсерции. Эта повторяющаяся последовательность ДНК, окаймляющая инсерцию, содержит обычно от 5 до 9 нуклеотидов.
Инсерционные последовательности относительно невелики и кодируют лишь функции, необходимые для их транспозиции. Второй класс подвижных элементов, так называемые транспозоны, содержит гены, не имеющие отношения к транспозиции, но придающие важные свойства клеткам бактерии-хозяина.
Впервые транспозоны были обнаружены, когда оказалось, что некоторые гены устойчивости к антибиотикам связаны с инфекционными факторами устойчивости. Отдельные гены устойчивости могут переноситься в другие плазмиды и хромосомы бактерий, почему и возник термин «транспозон».
Одна из полезных функций транспозонов состоит в том, что они способствуют включению в геном организмов новых, «чужих» генов. Другая возможная функция подвижных элементов может быть связана с их способностью вызывать различные хромосомные перестройки, в частности, соседние делеции и инверсии. Это может быть важным механизмом создания внутривидовой изменчивости хромосомных структур.
Эффект положения. Транспозиции и инсерции иногда активизируют близлежащий ген, приводят к изменению фенотипа вследствие того, что переместившиеся гены оказываются в новом окружении, а иногда изменяется и сам переместившейся ген. Это явление называется эффектом положения. Эффект положения может быть стабильным и нестабильным, или мозаичным.
Геномные мутации будут рассмотрены в следующей главе.
Вопросы для обсуждения
1. Что такое модификационная изменчивость и каково ее значение?
2. Что такое спонтанный мутационный процесс и каковы его закономерности?
3. Что такое индуцированный мутационный процесс и каковы его закономерности?
4. Что подразумевают под частотой мутации?
5. Какие мутации считают прямыми, а какие – обратными?
6. В чем различие между соматическими мутациями и мутациями в половых клетках? Доминантными и рецессивными мутациями?
7. Иногда точечные мутации называют генными. Как Вы полагаете, это допустимо? Почему?
8. Объясните, какая связь существует между мутациями и генами? Мутациями и рекомбинациями?
9. Когда хромосома разорвана, лежит ли точка разрыва внутри гена, между генами или возможны оба случая? Постарайтесь обосновать свой ответ.
10. В чем сущность закона гомологических рядов наследственной изменчивости? Назовите автора этого закона.
11. Объясните термины: нехватка, дупликация, инверсия, транслокация. Как возникают эти хромосомные нарушения?
12. Что такое «транспозоны», или «прыгающие гены»?
13. Объясните сущность «эффекта положения гена».
14. Насколько постоянным должно быть изменение в нуклеотиде, чтобы его можно было считать мутационным?
15. Замена каких азотистых оснований приводит к изменениям типа транзиции и трансверсии?
16. Нарисуйте как можно больше различных результатов разрывов хромосомы АВ/CDE/FGHJ/J, где точка обозначает центромеру, а наклонные линии – места трех одновременно произошедших разрывов. Укажите, какой из результатов должен встречаться чаще всего.
17. Как возникают кольцевые хромосомы?
18. Хромосома А.ВСDEEDCFG имеет перевернутую дупликацию участка CDE. Сравните устойчивость этой хромосомы с устойчивостью хромосомы A.BCDECDEFG, в которой есть смежная дупликация того же самого участка.
19. Изобразите конъюгацию хромосом, имеющих следующие расположения генов: ABCDEFGHM и AEDCBFHGM. Какие события могут предшествовать превращению первой последовательности генов во вторую?
20. Особь 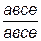 скрещивают с особью
скрещивают с особью 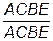 . Какиеё полноценные и дефективные продукты мейоза будет образовывать потомство этой пары?
. Какиеё полноценные и дефективные продукты мейоза будет образовывать потомство этой пары?
21. Как выглядит картина конъюгации в мейозе двух гомологичных хромосом, если одна из них утратила часть генов? Например, АВС.DЕКМNОР и АВС.DNОР.
22. Нарисуйте схему конъюгации хромосом в мейозе:
 К К D е
К К D е
е d Р Р
Дата добавления: 2015-01-02; просмотров: 13341;