Глава 12. ЭНДОКРИННАЯ СИСТЕМА РЫБ
Эндокринная система - одна из составляющих морфологической основы нейрогуморальной регуляции, прошедшей длинный эволюционный путь. В этой сложной системе гуморальная регуляция (в исто-рическом плане) первична. Система гуморальной регуляции развивалась параллельно с развитием клеточной мембраны, т. е. эта система регуляции имеется уже у одноклеточных животных. Примитивные одноклеточные обладают раздражимостью. Их клеточная мембрана проявляет избирательность, т.е. свойства, в основе которых функционируют гуморальные агенты- электролиты (ионы калия, натрия, хлора, кальция, водорода, гидроксила). Более сложные функции клетки (размножение, регенерация, рост) контролируются уже с участием гормонов или гормоноподобных веществ.
У многоклеточных организмов роль гуморальной системы в регуляции физиологических функций дополняется нервной системой, которая обеспечивает более адекватную реакцию животного на внешние и внутренние раздражители. Нервная и гуморальная системы функционируют как единое целое. Взаимодействие между ними осуществляется на трех уровнях: клеточном, органном, организменном.
Примером взаимодействия на клеточном уровне может служить работа нейрохимического синапса (рис. 12.1).
Нервный импульс поступает к регулируемой клетке по аксону соответствующего нейрона. Однако синаптическую щель нервный импульс преодолеть не может.
С этого момента функцию регуляции берет на себя гуморальное звено: под действием нервного импульса в пресинаптической мембране образуется химическое вещество - медиатор (например, норадреналин), который выбрасывается в синаптическую Щель, связывается рецепторами клетки-эффектора и вызывает соответствующие физиологические изменения в этой клетке.
Межорганное взаимодействие иллюстрирует работа гипоталамо-гипофизарно-адреналовой системы (рис. 12.2).
Афферентация рецепторного аппарата приводит в возбуждение определенные структуры таламуса. В результате возбуждается гипоталамус, а затем гипофиз. Здесь заканчивается нервное звено и начинается химическая регуляция.
Гипофиз вырабатывает и выделяет в кровь адренокортикотроп-гормон (АКТГ), который возбуждает интерреналовые тела головной почки рыб. Результатом этого возбуждения являются образование и выделение в кровь кортикостероидов.
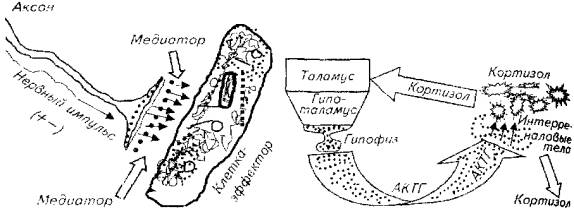
Рис. 12.1. Схема нейрохимического синапса
Рис. 12.2. Взаимодействие нервного и эндокринного (гуморального) звеньев из органном уровне
Примером взаимодействия нервной и гуморальной систем на организменном уровне является реакция адаптации, которая будет детально рассмотрена на рыбах ниже.
Главными агентами гуморальной регуляции служат гормоны, вырабатываемые специализированными клетками, которые могут быть вкраплены в слизистую оболочку, например, желудка, концентрироваться группами, образуя "тельца", например интерреналовые, или могут образовывать самостоятельные структуры, как в случае с щитовидной железой у млекопитающих. У рыб истинных желез внутренней секреции, кроме гипофиза, по существу, нет.
И тем не менее гуморальная система рыб имеет много общего с эндокринной системой высших позвоночных. Прежде всего эта общность затрагивает гормоны, многие из которых, например инсулин, тироксин, адреналин, не имеют видовой специфичности.
§55. ХИМИЧЕСКАЯ ПРИРОДА И МЕХАНИЗМ ДЕЙСТВИЯ ГОРМОНОВ
Химическая природа гормонов различна. Это и стероиды, и пептиды, и производные аминокислот, и производные жирны кислот. При этом все они обладают рядом общих черт:
контролируют синтез белка;
активны в чрезвычайно малых количествах;
имеют дистантный характер действия;
действуют только на свои клетки-мишени, имеющие реиепторы к конкретному гормону;
имеют непродолжительный период активности;
многие из них не имеют видовой специфичности.
Стероидные гормоны (кортизол, тестостерон, эстрадиол, прогестерон и др.) образуются из холестерина. К этой же группе специалисты причисляют арахидоновую кислоту и ее производные (простагландины, простациклины, тромбоксаны, лейкотриены).
Все стероидные гормоны гидрофобны; их транспортирование 0о кровеносному руслу осуществляют специальные переносчики. Однако внутрь клетки ввиду своей липофильности они проникают легко (рис. 12.3).
Рецепторы этих гормонов обнаруживают в цитоплазме клеток-мишеней. Белки-рецепторы одновременно выступают и транспортным средством в пределах клетки, доставляя гормон в клеточное ядро.
В ядре стероиды взаимодействуют с ДНК и вызывают синтез матричной РНК, затем рибосомальной РНК и полисом - комплекса дополнительных рибосом с мембранами эндоштазматического ретикулума. В результате индуцированной стероидным гормоном транскрипции и трансляции в клетке-мишени в течение нескольких часов образуется 3-5 новых белков.
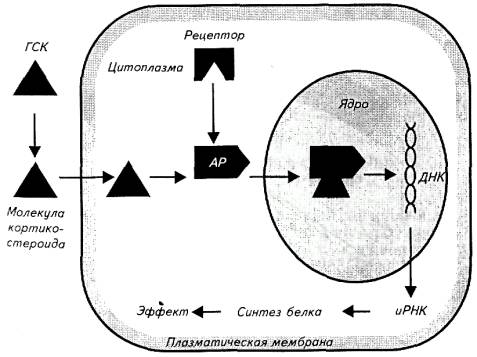
Рис. 12.3. Механизм действия стероидных гормонов:
комплекс стероидного гормона и молекулы глобулина крови; АР - активированный клеточный рецептор
Белковые гормоны (СТГ, ТТГ, ФСГ, ЛГ, пролактин, инсулин и др.) имеют слишком крупные размеры молекулы и не в состоянии самостоятельно проникать через клеточную мембрану (рис. 12.3) На поверхности клетки-мишени гормон улавливается рецептором. Механизм действия белковых гормонов заключается в активации цитозольных протеинкиназ, запускающих реакции синтез определенных белков.
Этому предшествует этап синтеза ряда химических веществ называемых месенджерами, под влиянием комплекса из гормона и белка-рецептора мембраны клетки-мишени. Роль месенджеров выполняют в основном три агента - цАМФ, ионизированный кальций и диацилглицерин.
Месенджеры активируют протеинкиназы, которые, в свою очередь, вызывают фосфорилирование белков и определенные физиологические эффекты (изменение проницаемости мембран, синтетические процессы, механические эффекты и др.).
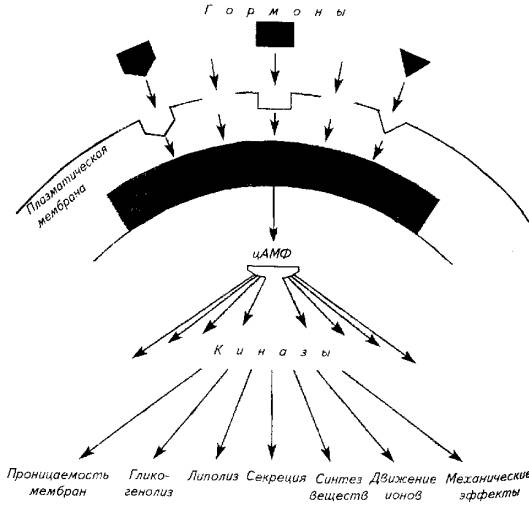
Рис. 12.4. Механизм действия гормонов нестероидной природы
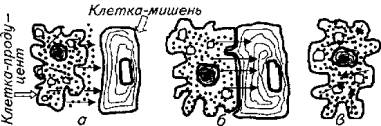
Рис. 12.5. Разновидности паракринной секреции:
а- паракринная; б - изокринная; в - аутокринная
цАМФ-зависимыми гормонами являются АКЛТ, ТТГ, ФСГ,
ЛГ, адреналин (через р-рецепторы). Кальцийзависимыми считаются окситоцин, вазопрессин, гастрин, катехоламины (через а-рецепторы).
Таким образом, эндокринной системе рыб больше подходит название "паракринная система", так как истинной железой можно считать только гипофиз. Остальные структуры, производящие гормоны, либо относятся к железам смешанного типа, либо являются паракринным аппаратом в чистом виде.
Различают несколько видов паракринных эффектов (рис. 12.5):
собственно паракринный механизм: специализированная клетка вырабатывает гормон, который поступает в межклеточное пространство и находит поблизости клетку-мишень;
изокринный механизм: клетка-продуцент гормона плотно контактирует с клеткой-мишенью, поэтому гормон переходит из клетки в клетку;
аутокринный механизм: клетка-продуцент одновременно является и клеткой-мишенью.
§56. ХАРАКТЕРИСТИКА ОСНОВНЫХ ЭНДОКРИННЫХ СТРУКТУР
Гипофиз справедливо считается главной железой внутренней секреции, поскольку прямо или опосредованно все железы зависят от него. В отличие от гипофиза высших животных гипофиз Рыб имеет упрощенное строение: он не имеет четко разграниченных передней, задней и средней долей (рис. 12.6).
Передняя доля гипофиза (аденогипофиз) у рыб может занимать большую часть железы. В этом случае его подразделяют на проаденогипофиз и мезоаденогипофиз.
Задняя доля гипофиза является продолжением промежуточного мозга, поэтому и называется нейрогипофизом. По существу, нейрогипофиз и не образует компактной доли у рыб. Чаще он имеет древовидную структуру и пронизывает своими ветвями всю массу гипофиза.
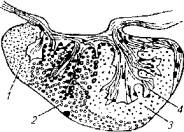
Рис. 12.6. Гипофиз костистых рыб:
1, 2- аденогипофиз; 3- промежуточная доля. 4- нейрогипофиз
Филогенетические корни аденогипофиза и нейрогипофиза различны. Аденогипофиз берет свое начало от органов пищеварения. В эмбриогенезе он формируется как выпячивание задней стенки глотки. Поэтому аденогипофиз рыб состоит из эпителиальной железистой ткани. Нейрогипофиз исторически связан с нервной трубкой. Поэтому он состоит из элементов нервной ткани - глиальных клеток, отростков нейро-секреторных клеток гипоталамуса и тел Герринга (особые окончания нейронов, выполняющих функцию накопителей нейросекретов). Средняя доля гипофиза у рыб отсутствует. Однако в составе гипофиза имеются скопления клеток, которые у рыб принято называть метааденогипофизом, являющимся аналогом средней доли гипофиза высших позвоночных животных.
Из гипофиза у рыб выделен тот же набор гормонов, что и у высших животных. Однако их физиологические эффекты на рыбе несколько отличаются от известных эффектов у высших позвоночных животных (рис. 12.7).
Соматотропный гормон (СТГ) образуется в передней доле гипофиза. Имеет различные органы-мишени. Основные физиологические эффекты на рыбе - рост, органогенез, регенерация органов и тканей.
Тиреотропный гормон (ТТГ) активизирует функцию щитовидной железы,
Адренокортикотропный гормон (АКТГ) контролирует активность интерреналовых тел головной почки. Является участником гуморальной фазы реакции адаптации при воздействии на организм стресс-факторов.
Пролактин имеет клетки-мишени в ряде органов: почках, жабрах, желудочно-кишечном тракте, плавательном пузыре, коже. Играет большую роль в регуляции избирательных свойств мембраны органов, причастных к осморегуляции у пресноводных видов рыб. Этот гормон стимулирует также образование кожной слизи.
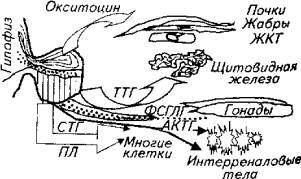
Рис. 12.7. Эффекты гормонов гипофиза у рыб
(ФСГ, ЛГ) гипофиза регулируют сперматогенез у самцов и овогенез у самок рыб. Влияют на половое поведение.
Окситоцин (ихтиотоцин) - гормон нейрогипофиза. Строго говоря, он вырабатывается гипоталамусом, а в нейрогипофизе просто накапливается. Этот гормон у рыб влияет на избирательную проницаемость клеток мембраны почек и жабр. Поэтому можно говорить о его причастности к осморегуляции. Интимный механизм его действия на рыбах изучен недостаточно.
Вазопрессин (вазотоцин) нейрогипофиза рыб регулирует тонус кровеносных сосудов и влияет на их проницаемость, регулирует водно-солевой обмен.
Меланофорный гормон (МФГ) метааденогипофиза у рыб, как и у других животных, регулирует пигментный обмен. Не все исследователи связывают происхождение этого гормона у рыб со средней долей гипофиза и причисляют его к продуктам нейрогипофиза. Однако, скорее всего, просто технические сложности не позволяют отделить продукт метааденогипофиза от продуктов нейрогипофиза, Секреторная активность метааденогипофиза стимулируется освещением. Выброс в кровь меланофорного гормона приводит к возникновению более темной окраски кожи рыб.
Щитовидная железа у рыб не оправдывает своего названия, заимствованного у высших позвоночных. У рыб под "щитовидной железой" понимают небольшое скопление специфических фолликулов на аорте между сердцем и жабрами и частично на мышцах нижней челюсти, Таким образом, собственно железы как таковой у большинства костистых рыб нет (рис. 12.8).
По некоторым данным, тиреоидные фолликулы встречаются и на стенках спинной аорты, в головной почке и даже в ткани селезенки. У двоякодышащих и, как ни странно, у хрящевых рыб тиреоидные фолликулы формируют компактный орган.
Из тиреоидных образований рыб выделены три гормона - Т2, ТЗ и Т4. Из них в крови обнаруживается тетраиодтиренин (14). Функция тиреоидных гормонов у рыб до конца неясна. Тироксин не дает у рыб такого однозначного эффекта по отношению к основному обмену, как у наземных животных. Метаморфоз, органогенез и регенерация тканей у рыб зависимы от тиреоидных гормонов лишь частично. Исследователи отмечают гиперфункцию тиреоидных фолликулов у рыб при сезонных, пищевых миграциях и при нересте.
Эвригалинные рыбы при попадании в соленую воду также отвечают повышением функциональной активности щитовидной железы, что на-
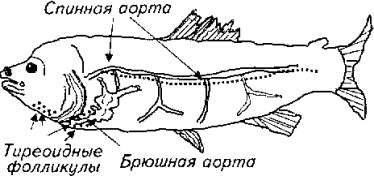
Рис. 12.8. Топография тиреоидных образований у рыб ("щитовидная железа")
водит на мысль о причастности тиреоидных гормонов к осморегуляции. Есть свидетельства того, что пероральное назначение тиреоидных гормонов вызываег усиление роста молоди рыб.
Иптерреналовые тельца (функциональный аналог корковой части надпочечников) невооруженным глазом увидеть невозможно так как они представляют собой микроскопические вкрапления в ткань головной почки. Кортикостероиды рыб аналогичны и по химическому составу, и по физиологическим эффектам гормонам корковой части надпочечников высших позвоночных. Из глюкокортикоидов рыб наиболее изученными являются кортизол и кортикостерон.
Кортизол обеспечивает адаптацию организма рыб в стрессовых ситуациях. Он стимулирует глюконеогенез в печени, поддерживает на постоянном уровне концентрацию глюкозы в крови, подавляет жироотложение. Этот стероид у рыб причастен и к процессу осморегуляции в пресной воде, так как активно удерживает натрий в почечных канальцах.
Кортикостерон тоже участвует в осморегуляции. Поскольку он выражение подавляет гломерулярную фильтрацию воды, то можно с уверенностью говорить об его особой роли в процессе поддержания водно-солевого гомеостаза у рыб в соленой воде.
Роль минералокортикоидов у рыб изучена крайне плохо. Известно, что главный минералокортикоид млекопитающих - альдостерон - на рыбах неэффективен.
Хромаффиновые железы - аналоги мозгового слоя надпочечников - секретируют в кровь катехоламины. Хромаффиновые клетки можно обнаружить при гистологических исследованиях головной почки в области ее прилегания к задней кардинальной вене. При помощи гистохимических методов в цитоплазме хромаффиновых клеток выявляют два гормона - норадреналин и адреналин. Есть сообщения о том, что соотношение этих гормонов в крови рыб иное, чем у наземных животных. По крайней мере, у карпа преобладающей формой является норадреналин.
Физиологические эффекты катехоламинов на рыбах аналогичны таковым наземных животных, т. е. они дублируют симпатические влияния. В стресс-реакциях на стадии тревоги у рыб в крови повышается уровень адреналина и возникают классические изменения физиологических показателей.
Ренин-ангиотензиновая система у рыб представлена окологломелулярными клетками, т. е. является еще одной эндокринной системой в составе почек. Гормон этой структуры -ренин - довольно хорошо изучен. Его основная роль - активация ангиотензина-1 (вырабатывается печенью) в ангиотензин-Н, повышающий кровяное давление в почках. Ренин является видоспецифичным гормоном. Выделенный из почек рыб, он у млекопитающих не вызывает прессорных реакций. Ренин образуется и в тельцах Станниуса - еще одном почечном образовании рыб.
Тельца Станниуса - это два микроскопических образования на цетральной стороне каудальной части почки рыб. Гистохимические исследования, к сожалению, не позволяют выделить в их структуре какой-либо из известных у млекопитающих гормон или гормоноподобное вещество. Однако мало кто сомневается в их эндокринном предназначении, так как их экстирпация сопровождается эффектом выпадения с явными признаками нарушений гормональных регуляций. Их удаление у рыб вызывает серьезные нарушения водно-солевого гомеостаза. Некоторые исследователи считают, что тельца Станниуса вырабатывают вещество гипокальцин, влияющее на обмен кальция у рыб.
Улътимобранхиальная железа - структура, обнаруженная на мембране между сердцем и печенью, имеет вид белесой полоски. Клетки, ее образующие, секретируют в кровь гормон кальцитоцин. Этот гормон вызывает сильнейший гипокальциемический эффект у человека и, что совершенно неожиданно, не влияет на уровень кальция в крови у лососей, из железы которых он был выделен. Объяснить это странное явление, наверное, можно следующим. У рыб кальциемию контролирует несколько систем. Роль кальцитонина, возможно, сводится только к контролю за экскрецией кальция через жабры при содержании рыб в соленой во-де. В пресной воде его жаберно-экскреторная функция блокируется.
Урофиз - нейросекреторная зона концевого отдела спинного мозга - был обнаружен при изучении гистоструктуры спинного мозга еще в середине XIX века. Однако эта железа остается загадкой для физиологов и по сей день.
Установлено, что урофиз вырабатывает и секретирует в кровь несколько близких по химическому строению веществ. Их называют уротензинами. У карпа эта железа вырабатывает также ацетилхолин. Считается, что уротензины участвуют в осморегуляции пресноводных рыб. Экспериментально доказано, что они снижают периферическое давление крови, вызывают сокращения гладких мышц и способствуют сохранению натрия в организме пресноводных рыб. Уротензины стимулируют поглощение натрия из воды жаберными мембранами и такое же поглощение натрия в почках из первичной мочи.
Поджелудочная железа - железа смешанного типа. Она представляет собой компактный орган только у хрящевых рыб. У костистых она объединена с печенью и потому называется гепатопанкреасом. Этот орган располагается вдоль кишки и достигает разной степени развития, составляя от 1,5 % живой массы у карпа до 10% живой массы у трески.
Разбросанные по печеночной паренхиме дольки поджелудочной железы имеют индивидуальные эк-зосекреторные протоки, которые после слияния впадают в желчный пузырь. У некоторых видов экзосекреторная часть поджелудочной железы имеет собственные протоки, открывающиеся в головной кишке и пи лирических придатках.
Эндокринная часть разбросана по долькам железы и формируется из клеток четырех типов, включая нервные клетки. Как и у высших позвоночных, поджелудочная железа как эндокринный орган причастна к регулированию углеводно-жирового обмена. Однако в экспериментах после парентерального введения инсулина далеко не всегда получают гипогликемический ответ.
Вероятно, глюкоза крови не является для рыб жесткой константой гомеостаза, так как в норме ее концентрация в крови колеблется от 50 до 150 мг%. Более того, экспериментально у рыб удается довести концентрацию глюкозы в крови до нуля без каких-либо клинических признаков патологий, например судорог (как у млекопитающих). Объяснить это можно тем, что у рыб глюкоза не является главным источником энергии для мозга и мышц, как у наземных животных. В мышечном энергетическом обмене у рыб основную роль источников энергии выполняют липиды. И тем не менее во многих случаях инсулин понижает уровень глюкозы в крови рыб, а его антагонист глюкагон соответственно вызывает противоположную реакцию.
Гонады как железы смешанной секреции имеют хорошо развитый эндосекреторный аппарат, который вырабатывает половые стероиды.
В ястыках вырабатываются в больших количествах эстрогены, которые регулируют у самок овогенез, влияют на развитие вторичных половых признаков при половом созревании, формируют половое поведение самок.
Определяющая роль эстрогенов в половом детерминизме ювенальных гермафродитов доказана в экспериментах по скармливанию эстрона и эстрадиола молоди тиляпии и лососевых рыб. Причем эстрадиол оказывал более сильный по сравнению с эстроном эффект. При его использовании и на тиляпии, и на лососях выход самок составлял 100 %.
Роль прогестерона у самок рыб пока изучена плохо.
Активность эндокринной системы яичников характеризуется цикличностью. Большую часть года она находится в состоянии относительного покоя. С приближением нерестового периода под влиянием гонадотропных гормонов гипофиза происходит ее активизация.
Семенники рыб вырабатывают мужские половые гормоны, наибольшее значение из которых имеет тестостерон. Андрогены стимулируют сперматогенез, формируют вторичные половые признаки, опреде-ляют половое поведение самцов при нересте. Тестостерон и другие мужские половые гормоны обладают ярко выраженным анаболическим эффектом и при добавлении в корм уже в количестве 1 -10мг/кг вызывают ускоренный рост многих видов рыб (карась, форель, тиляпия, кижуч).
Таким образом, эндокринная (паракринная) система рыб регулирует основные вегетативные функции рыб и определяет многие стереотипические поведенческие реакции.
Глава 13 ИММУНИТЕТ У РЫБ
Под иммунитетом понимают способность организма к самозащите от чужеродных корпускулярных биологических тел, а также веществ, несущих чужеродную генетическую информацию.
Иммунная система рыб представлена нефросом, селезенкой, лимфоидным органом, печенью, а также множественными включениями лимфоидной ткани в желудочно-кишечном тракте, сердце, жабрах. Непосредственными исполнителями функции защиты являются органы с барьерными функциями - кожа, жабры, желудочно-кишечный тракт, а также лейкоциты (все виды).
Различают специфический и неспецифический иммунитет.
Специфический иммунитет обеспечивает защиту организма от строго определенных антигенов. По существу, иммунная реакция организма сводится к уничтожению, нейтрализации или удалению из организма антигена.
Неспецифический иммунитет осуществляет защиту организма от любого чужеродного вещества или тела.
Механизм элиминации антигена может быть клеточным или гуморальным.
Клеточный иммунитет предполагает прямую атаку антигена клетками иммунной системы, например фагоцитоз или пиноцитоз.
Лейкоциты могут элиминировать антиген и на расстоянии, не вступая с ним в прямой контакт. Это явление называется цитотоксинеским эффектом.
Гуморальный иммунитет обеспечивают различные структуры как иммунной системы, так и других систем. Он заключается в продуцировании различных химических веществ - лизопима, интерферона, кининов и других веществ, слизи, антител, которые так или иначе инактивируют антиген.
Неспецифический иммунитет является врожденным свойством организма. Его клеточную форму обеспечивают нейтрофилы, моноциты, макрофаги, клетки РЭС (рис. 13.1).
Гуморальная форма неспецифического иммунитета обеспечивается различными органами и клетками, производящими лизоцим, интерферон, пропердин, комплемент, фибронектин и некоторые другие биологически активные вещества. Часто общую оценку неспецифического иммунитета проводят по активности лизоцима сыворотки крови. Этот фермент с ярко выраженными бактерицидными свойствами вырабатывается в организме всех позвоночных. Сообщения о его отсутствии у того или иного вида рыб, скорее всего, связаны с некорректными оценками его активности. У рыб сезон года, физиологическое состояние сильно меняют титр лизоцима. Так, у карпа активность лизоцима максимальна осенью, в конце сезона выращивания, и минимальна в конце зимовки. Есть сообщения, что хищные рыбы имеют более высокие титры лизоцима по сравнению с мирными рыбами.
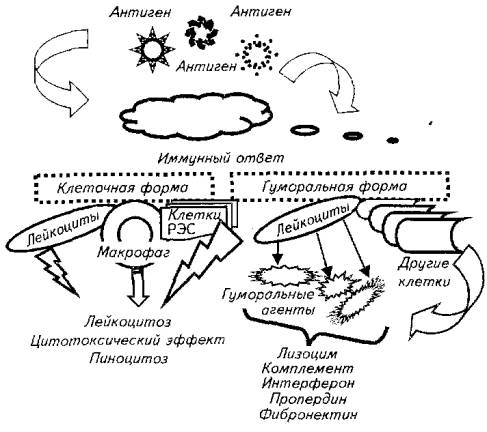
Рис. 13.1. Схема неспепифического иммунного ответа
Интерферон - группа веществ, происхождение которых связывают с макрофагами, нарушает процесс размножения вирусов. Интерферон обнаруживается в крови всех рыб, но у мирных видов его титры выше, чем у хищников.
Комплемент - группа белков (около 20) - продукт жизнедеятельности макрофагов. Он способен обезвреживать как корпускулярные, так и гуморальные чужеродные вещества. Комплементу приписывают несколько функций:
опсонизация - взаимодействие с чужеродной клеткой с целью ее подготовки к фагоцитозу;
цитолиз - цитотоксическое воздействие на антигены клеточного типа;
активизация тучных клеток и привлечение фагоцитов в очаг воспаления;
нейтрализация антигенов и дальнейшее их удаление из организма;
Фибронектин - еще один гуморальный компонент неспецифического иммунитета. Он вырабатывается во многих клетках, включая макрофаги. Механизм его действия напоминает комплементарную опсонизацию.
Пропердин - бактерицидное вещество, вырабатываемое моноцитами и макрофагами. В большом количестве присутствует в крови карповых и других рыб.
Кинины - мелкие полипептиды с разносторонними функциями. Они вызывают сокращение и расслабление гладких мышц повышают сосудистую проницаемость (в первую очередь посткапиллярных венул), активируют фактор Хагемана системы коагуляции. Кинины в большом количестве содержатся в кожной слизи рыб.
Клеточный неспецифический ответ обеспечивают в основном лейкоциты посредством следующих механизмов:
фагоцитоз;
пиноцитоз;
цитотоксический эффект;
синтез гистамина, серотонина, простагландинов;
удержание антигена на пути в кровеносную систему.
Фагоцитоз характерен для нейтрофилов, одна клетка которых способна поглотить десятки корпускулярных тел. Нейтрофилы вырабатывают активные соединения кислорода - перекись водорода, супероксидный радикал, которые оказывают разрушительное влияние на клеточные мембраны микробов. Лизосомальные ферменты нейтрофилов растворяют микробные клетки. Подобным образом действуют и другие фагоцитирующие клетки - моноциты, макрофаги.
У рыб в определенных условиях отмечают фагоцитоз у эритроцитов, тромбоцитов, клеток РЭС, некоторых клеток слизистой оболочки желудочно-кишечного тракта.
Пиноцитоз напоминает фагоцитоз, но происходит за счет инвагинации клеточной мембраны, которая как бы засасывает антиген внутрь цитоплазмы. Пиноцитозу подвергаются не корпускулярные жидкие чужеродные вещества.
Цитотоксический эффект осуществляется как при непосредственном контакте с антигеном, так и на расстоянии за счет атаки клетки-мишени перекисью водорода, гипохлорной кислотой. Цитотоксический эффект свойствен в той или иной степени всем видам лейкоцитов и макрофагам.
Специфический иммунитет (рис. 13.2) возникает при обязательном участии лимфоцитов и имеет два варианта развития: клеточный и гуморальный.
Клеточный ответ осуществляют Т-лимфоциты-киллеры. После активации Т-киллера чужеродным антигеном он убивает клетку-мишень (бактерию, вирус, гриб) цитотоксическим способом (супероксидный ион, перекись водорода, лимфотоксин), вступая или не вступая с ней в непосредственный контакт.
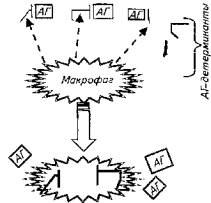
Рис. 13.2. Схема процессинга
Гуморальный ответ лимфоцитов выглядит сложнее. Для удобства его изучения можно разбить этот процесс на три этапа:
распознавание антигена и активация исполнительных клеток;
вовлечение клеток памяти и исполнителей элиминации антигена;
процесс элиминации антигена. В качестве антигенов животный организм воспринимает такие вещества, как белки, полисахариды, нуклеиновые кислоты, в виде отдельных молекул с большой массой или групп молекул или корпускулярные тела. Независимо от химической природы и сложности строения любой антиген имеет специфические функциональные группы - антигенную детерминанту, количество которых определяет валентность антигена. Для запуска иммунного ответа необходимо, чтобы в организм попал определенный минимум единиц антигена. Антиген улавливается еще в тканях организма макрофагами, но если все же антиген успевает попасть в кровяное русло, то он атакуется Т-лимфоцитами-хелперами, которые собирают антигены в "кассету".
Макрофаги подвергают антигены процессингу, т. е. концентрации антигенных детерминант (см. рис. 13.2). За счет процессинга антигенность чужеродного тела или вещества возрастает в десятки и сотни раз.
После процессинга макрофагами антиген передается Т-лимфоциту-хелперу, который, в свою оче-редь, предоставляет его для элиминации или Т-лимфоцитам-киллерам, или В-лимфоцитам для формирования гуморального ответа (рис. 13.3).
Последний процесс получил название кооперации Т- и В-лимфоцитов. Он не так прост и состоит из трех фаз.
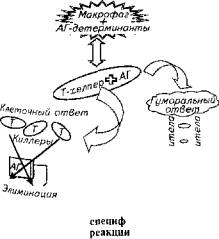
Рис. 13.3. Схема специфической имунной реакци
1. Распознавание антигенной детерминанты В-лимфоцитами. Каждый лимфоцит имеет на своей клеточной мембране десятки тысяч молекул у-глобулинов, которые выполняют функцию рецептора к антигену. Поэтому один В-лимфоцит распознает сотни тысяч антигенов разной химической природы и организации.
2. Т-лимфоцит-хелпер отыскивает комплекс макрофаг - антиген, после чего начинает продуцировать интерлейкин-II, т. е. вещество, которое инициирует трансформацию части В-лимфоцитов в плазматиче-скую клетку - продуцент антител.
3. Макрофаг в ответ на действия t-хелпера секретирует интерлейцин-1. Последний стимулирует усиление образования В-лимфоцитов лимфоидными органами для компенсации трансформированных в плазматические клетки В-лимфоцитов крови. Адекватность компенсаторного ответа лимфоидной ткани находится под контролем Т-лимфоцитов-супрессоров.
Таким образом, агентом гуморального специфического ответа выступает антитело к данному антигену, продуцируемое трансформированным В-лимфоцитом (рис. 13.4). По химической природе все антитела -гликопротеиды. В составе антитела имеется особая зона, определяющая специфичность данного антитела. Благодаря этой зоне антитело узнает "свой" антиген.
У млекопитающих выделены антитела пяти классов (иммуноглобулины G, M, A, D, Е), которые от-личаются первичной и вторичной структурой. У рыб удается найти только два из них: IgG и IgM. Элиминация антигена при воздействии на него антитела происходит путем:
преципитации растворимых в сыворотке антигенов; агглютинации чужеродных клеточных структур;
лизиса свободных от комплемента антигенов.
Кроме того, антиген подвергается нейтрализации путем связывания с антителом. Возможна и опсонизация антигена, т. е. образование комплекса, который легко фагоцитируется макрофагом. Собственно защита организма от специфического антигена возникает тогда, когда в сыворотке крови лимфоцитами вырабатывается определенное количество антител. Это возможно в двух случаях.
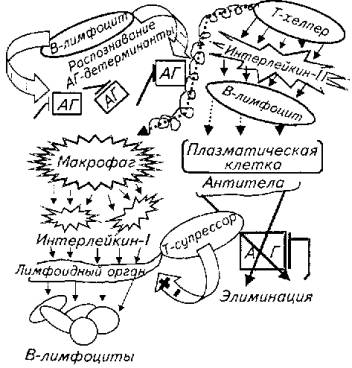
Рис. 13.4. Гуморальный ответ специфического иммунитета
Во-первых, когда антиген попадает в организм животного естественным путем в достаточном для запуска ответа количестве. В этом случае формируется естественный иммунитет.
Во-вторых, образование антител можно спровоцировать, например, введением ослабленной культуры микроорганизмов.
В этом случае иммунный ответ развивается менее болезненно тля организма животного. Однако эффективность искусственного иммунитета ненамного уступает естественному иммунитету.
Возможно введение готовых антител в кровь (пассивный иммунитет), однако при этом, как правило, продолжительность иммунной защиты невелика.
Искусственная иммунизация против инфекционных заболеваний применяется и в рыбоводстве. Основным методом иммунизации является включение в корм рыбы ослабленных форм антигенов. В условиях товарного производства рыбы парентеральная иммунизация по экономическим соображениям невозможна.
Пассивная иммунизация (введение сыворотки с антителами) используется в редких случаях, чаще в экспериментах, для сохранения ценных видов рыбы (например, для борьбы с вибриозом лососевых).
Глава 14 СТРЕСС У РЫБ
Стресс у высших животных и человека - это прежде всего психофизиологическая реакция, в формировании которой важнейшее место занимает кора больших полушарий.
У рыб психоэмоциональных состояний такого рода нет либо они находятся в зачаточном состоянии, так как у них нет не только коры, но и больших полушарий (исключение составляют некоторые виды двоякодышащих рыб).
Из рис. 14.1 видно, что у рыб сенсорные системы и их центры обладают высокой степенью развития и обособленности.
В классе рыб отмечается большое разнообразие морфологических особенностей головного мозга. Причем в строении головного мозга рыб не просматривается связи между эволюционным положением вида и организацией центральной нервной системы.
У разных видов рыб отдельные части головного мозга имеют неодинаковый уровень развития. Так, у активных видов акул (катран) мозжечок хорошо развит, а у их родственников (скатов) зачастую эту часть головного мозга даже обнаружить непросто. У карпа хорошо развиты обонятельные доли и обонятельные луковицы, а у кефали обращают на себя внимание сильно развитые зрительные доли. У карповых в заднем мозге имеется два крупных образования, напоминающих доли, - сильно развитые ядра нервов X пары (вагусные доли), а у других видов рыб они невооруженным глазом не видны.
Передний мозг у рыб (кроме двоякодышащих) ввиду примитивного строения не может выполнять интегрирующие функции, как у высших позвоночных. У рыб функцию интеграции выполняет лимбическая система, например промежуточный мозг, который и у млекопитающих, включая человека, играет далеко не последнюю роль в формировании психоэмоциональных состояний. Промежуточный мозг у рыб развит хорошо. Поэтому определение стресса как психофизиологической реакции организма вполне допустимо и по отношению к рыбе.
У рыб в качестве стрессорных факторов выступают абиотические факторы внешней среды (температура, химический состав воды, рН, содержание кислорода и др.), факторы внутренней среды (болевые ощущения, изменения констант внутренней среды, инфекционно-инвазионные интоксикации), социальные факторы (иерархическое положение, контакт с хищником, половым конкурентом), антропогенные факторы (контрольный облов, пересади
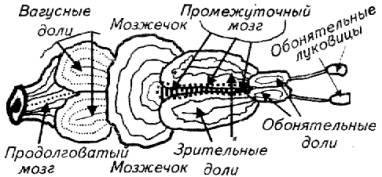
Рис. 14.1. Схема строения головного мозга рыбы
из зимовальных в нагульные пруды, транспортирование рыбы, ветеринарные мероприятия, высокие плотности посадки и др.).
Во всех перечисленных случаях наблюдается генерализованная реакция организма рыбы, которая сводится к адаптации к изменившимся условиям (восстановление констант внутренней среды, достижение полезно-приспособительного эффекта в соответствии с той или иной программой стереотипического поведения). При этом может быть два результата.
1. Если стрессор был в меру сильным, или действовал ограниченное время, или изменения среды происходили не очень быстро, то организму удается адаптироваться. Более того, в этом случае чаще всего стресс-фактор оказывает положительное влияние на организм рыбы.
2. Возможности рыбы к адаптации исчерпаны, а полезно-приспособительный эффект не достигнут.
В соответствии с классическими представлениями Г. Селье стресс имеет три стадии.
1. Стадия тревоги. Характеризуется резким усилением симпатического влияния на вегетативные и соматические реакции организма.
2. Стадия резистентности. Характеризуется устойчивым состоянием гомеостаза и поведенческих реакций.
3. Стадия истощения. Напоминает первую по напряжению симпатоадреналовой системы, которая развивается, однако, на фоне глубокого истощения организма. Поэтому на этой стадии возможна гибель рыбы.
Для рыб пока не разработано таких унифицированных показателей глубины стресса, как для млекопитающих (уровень катехоламинов и кортизола в крови, психоэмоциональные и вегетативные реакции). Механический перенос этих показателей на Представителей класса рыб невозможен по ряду причин. Так, например, не совсем понятен адреналовый ответ. У карпа катехоламины представляет главным образом но-радреналин (70 %), у кижуча - адреналин. Другие виды рыб выбрасывают равные количества адреналина и норадреналина в кровь. У форели уровень катехоламинов в крови в состоянии физиологического покоя со-ставляет примерно 100мкг%, а после получасового содержания без воды-20 000 мкг%.
По данным разных авторов, рост концентрации кортизола у Рыб отмечается не ранее чем через 15 мин после начала воздействия стресс-фактора. Однако количественная характеристик кортикального ответа рыб изучена недостаточно.
С некоторой натяжкой у рыб в качестве индикаторов глубины стресса используют концентрацию глюкозы и хлоридов в крови. Однако, принимая во внимание мягкость этих констант гомеостаза у рыб, вряд ли их можно признать универсальными показателями стрессированности рыбы.
Автор предпринял попытку увязать клинические показатели и локомоторные реакции рыб с глубиной стресса (рис. 14.2). В качестве стресс-фактора использовали рН воды (резкое изменение с 7,0 до 12,0), а в качестве критериев стрессированности рыбы - частоту дыхания, стайное и индивидуальное поведение, а также рефлексы позы. Опыты проводили на тиляпии живой массой около 50 г при ее содержании в бассейнах при температуре воды 28 њС. Наблюдения показали, что частота дыхания рыб коррелируете поведенческими реакциями, а на стадии истощения рыбы -и с органическими изменениями.
Стадия тревоги у стайных рыб проявляется очень ярко. Для этой стадии характерны особая сплоченность стаи и общая настороженность по отношению к любому раздражителю внешней среды-свету, звуку, появлению в бассейне инородного предмета. Стая при этом или держится у дна, или удаляется в угол бассейна. Частота движений жаберных крышек повышается по сравнению с состоянием покоя в два раза.
Не составляло большого труда определить и границу начала стадии истощения. Стая "рассыпалась", периодически рыба всплывала к поверхности и жадно хватала ртом воздух. Дыхание было частым и неритмичным. Рыба ложилась на бок, плавала кругами в таком положении или опускалась под углом 45њ. При длительном пребывании рыбы в воде с рН более 30,0 развивались органические изменения (повреждения слизистой глаза, кожи на грудных плавниках с последующим некрозом тканей).
При вскрытии рыб констатировали нарушение процесса желчеобразования - преобладание в со-ставе желчи билирубина вместо биливердина.
При достижении рН 12,0 часть рыб исчерпывала свои адаптационные возможности и погибала.
Сложнее было уловить границы стадии адаптации. Условно мы их определили в рН 8,5-10 рН. В этом диапазоне стая успокаивалась. Рыбы начинали пересматривать свое иерархическое положение, естественно, стремясь занять более высокое положение в "обществе". Восстанавливались пищевые рефлексы. Все это происходило на фоне нормализации частоты дыхания.
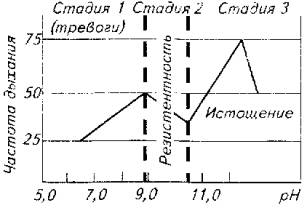
Рис. 14.2. изменения рН воды и глубина стрессового состояния тиляпии
Несомненно, более объективно можно судить о глубине стресса П0 комплексу клинических, поведенческих и биохимических показателей. Однако при работе с рыбой это не всегда возможно по техническим причинам. Поэтому в особых случаях при использовании рыб-аналогов, хорошем знании биологии вида границы стадий стресс-реакции можно определять только по поведению и доступным клиническим показателям.
Последние исследования показали, что адренокортикальный ответ гораздо сложнее, чем принято считать. Он возникает в том случае, когда симпатико-адреналовый механизм не обеспечивает достижения полезно-приспособительного эффекта. Центральным событием адренокортикального механизма является выделение глюкокортикоидов (кортизол, гидрокортизон). Эти гормоны повышают глюконеогенез, что восстанавливает уровень глюкозы и пула жирных кислот в крови, утраченных в результате симпатико-адреналовой накачки.
Избыточная продукция кортизола при затяжном воздействии стресс-факторов приводит к нежелательным последствиям, т. е. переводит стадию резистентности в стадию истощения. В этих условиях лейкоциты и лимфоидная система в целом атрофируются, так как белки, составляющие белые клетки крови и лимфоидные структуры, используются в процессе глюкоонеогенеза. Результатом являются резкое понижение защитных реакций организма и развитие вторичных патологий с инфекционным началом. Последнее находит благоприятную почву в стрессированном организме на фоне возникших геморрагических изменений (язвы желудочно-кишечного тракта, жаберного аппарата, плавников).
Современные представления о развитии стресс-реакции не ограничиваются симпатико-адреналовым ответом. Возбуждение гипоталамуса при воздействии стрессора приводит к выделению соматолиберина, т. е. активизирует соматотропный механизм. Одна из функции СТГ - противодействие инсулину, что способствует гомеостазу глюкозы и жирных кислот крови.
Также возникает и тиреоидный ответ. Выброс ТТГ активизирует тиреодные структуры. Тироксин повышает чувствительность клеток к действию катехоламинов и в целом повышает уровень основного обмена, что изменяет ряд клинических показателей - Частоту дыхания, сердцебиения, повышает артериальное давление.
Стрессовое возбуждение гипоталамуса стимулирует образование гипофизом Б-липотропина, являющегося предшественником так называемых эндогенных опиатов - энкефалина, эндорфина, инорфина. У млекопитающих опиаты снижают психогенную реактивность, т. е. способствуют адаптации к стресс-воздействию и переводят стадию тревоги в стадию резистентности (подавляют чувство тревоги, нормализуют соматические ответы).
Кроме опиатов на психогенную адаптацию влияют и другие системы. Среди них:
ГАМК-эргическая система. у-аминомасляная кислота (ГАМК) сама по себе является тормозным медиатором, следовательно способна подавлять избыточную стресс-реактивность. ГАМК в структурах центральной нервной системы, и прежде всего в промежуточном мозге, превращается в уокс и масляную кисло-ту (ГОМК). В экспериментах на высших позвоночных доказан сильнейший антистрессовый эффект ГОМК;
простагландин PgE снижает чувствительность нейронов к катехоламинам;
парасимпатическое влияние тропных ядер гипоталамуса и вагусных долей стимулирует восстановительные процессы и стабилизирует константы гомеостаза.
Таким образом, при стресс-ответе организма происходит конкуренция двух противоборствующих систем: 1) системы, мобилизующей энергетические ресурсы организма ради спасения жизни, но в ущерб иммунной и другим системам; 2) системы, ограничивающей глубину стресс-ответа. Сила и продолжительность стресс-воздействия существенно влияют на баланс сил этих систем.
Дата добавления: 2014-12-22; просмотров: 7348;